注:本文为 “群体选择与合作” 相关文章合辑。
人类合作之谜:群体选择及其演化仿真研究
刘业进 杨美荣
摘要:基因 - 文化共同进化塑造了人类物种。专业化和分工是人类独特的生存策略和行为模式,一系列先天心理倾向和文化本能等基础性适应器发展出来,通过组织和市场交换支持这种生存策略和行为模式。群体选择是驱动人类合作的主导力量。普莱斯方程揭示了群体间选择与群体内个体选择的净效应如何导致合作的进化。本文基于普莱斯方程的数值模拟和演化仿真发现,尽管非合作策略在 “群体内个体选择” 中占优,而在考虑 “群体间选择” 的净效应是合作策略占优。把合作行为收益 b 作为测度合作剩余的平均项,演化仿真发现,群体中合作者频率越高、平均适应度越大,都会导致所需 b 值降低,而当迭代次数越多,所需 b 值增加;合作者成本成倍增加时,所需最小 b 值是成倍增加;当加入促进合作的 “执行机制”(迭代后的合作者频率乘 1.001),所需 b 值降低。利己和利他作为主观行为动机在很多场合不具备第三方不可证实性,而合作和非合作则在人类实际生活和实验室实验中具有第三方可证实性,因此本文纠正传统上利己、利他这一对具有误导性的概念,提出将 “合作行为” 替换 “利他主义行为”,“非合作行为” 替换 “利己主义行为”,使群体选择理论在解释人类合作秩序的涌现时更具一般性。
关键词:普莱斯方程;群体选择;文化演化
DOI:10.19592/j.cnki.scje.400658
JEL 分类号:B52,C92,D71
中图分类号:C912
文献标识码:A
文章编号:1000 - 6249 (2022) 11 - 036 - 22
卢梭的 “猎鹿博弈” 和休谟的 “划船博弈”(表 1;表 2),说明分工和专业化在人类生存繁衍中的极端重要性。在猎鹿博弈中,猎鹿均衡有巨大的 “合作剩余”,但存在合作要求和风险;猎兔均衡不需要任何合作要求,且风险更小,但 “合作剩余” 小。划船博弈也存在两个类似的纳什均衡。群体选择是从 “猎兔均衡” 走向 “猎鹿均衡” 的进化动力。在猎鹿博弈和划船博弈中,“空间结构(局部互动)”、“信号机制” 和 “回报强化的互动动态” 是维持群体选择的三个决定因素。本文考察群体选择如何维持合作条件,属于 “回报强化的互动动态” 范畴。
表 1 猎鹿博弈

| 猎鹿 | 猎兔 | |
|---|---|---|
| 猎鹿 | (10,10) | (0,2) |
| 猎兔 | (2,0) | (2,2) |
表 2 划船博弈
| 合作 | 不合作 | |
|---|---|---|
| 合作 | (10,10) | (-2,0) |
| 不合作 | (0,-2) | (0,0) |
- 刘业进,首都经济贸易大学城市经济与公共管理学院,E-mail:[email protected],通讯地址:北京市丰台区张家路口 121 号首都经济贸易大学,邮编:100070;杨美荣(通讯作者),首都经济贸易大学城市经济与公共管理学院,E-mail:[email protected]。作者文责自负。
一、文献回顾
非血缘关系的个体间大规模合作在动物世界只见于人类。韦永贵、张艺川 (2021) 研究表明,跨文化传播显著推动了中国出口贸易的发展。这种 “人类合作秩序” 一直是演化经济学和其他相关学科关注的焦点,2005 年《Science》创刊 125 周年之际将其列为 25 个未解之谜之一。不同学科的学者对其进行了深入研究,学术界已经形成了一些相对成熟的理论,如:亲缘选择、直接互惠理论、间接互惠理论、文化群体选择理论 (CGS) 等 (Richerson et al,2016)。国内学术界的研究也逐渐增多。
黄少安、张苏 (2013) 将博弈思想用于解释人类合作以及演进,如果提供符合亲缘选择、直接互惠、强互惠、网络互惠、组间互惠的动机,纳什均衡可以逻辑一致地解释自然选择形成合作演化。而张衔、魏中许 (2016) 认为黄少安并没有破解 “人类合作之谜”,没有解释自然选择如何导致合作行为,后者认为 “人类合作之谜” 不完全是博弈论所揭示的 “策略合作博弈”,而更多的是博弈论不能解释的 “非策略性合作之谜”,并认为马克思的唯物史观和剩余价值理论揭示了人类合作的经济基础。
郭颖 (2011) 归纳认为人类合作有三种理论范式:神学范式、人性论范式和进化论范式,进化论范式解释人类合作本能的起源具有无可替代的优越性。赵阳、姜树广 (2015) 认为领导 - 追随模式是维持人类合作秩序的重要机制,领导通过惩罚机制和等级制统治可以克服搭便车问题。刘永芳、王修欣 (2019) 提出了有限理性合作观,包含了三个基本论断:合作决策是有限理性的启发式过程;有限理性启发式让人更注重合作;有限理性导致的合作行为具有一定的社会适应性。
韦倩等 (2019) 仿真结果表明,非协调性惩罚很难维持较大规模群体中合作的演化,协调性惩罚 (惩罚者相互配合实施的惩罚行为) 会提高群体合作水平。许多研究者质疑经典进化理论 (直接互惠、声誉理论等) 不足以解释人类合作,Krasow and Delton (2016) 结合心理学分析认为经典理论可以解释已知行为。Fehr and Rockenbach (2004) 表明强互惠、利他惩罚和利他奖励的行为倾向,对人类合作至关重要。Shutters (2012) 认为亲缘选择、直接互惠、间接互惠、多级选择等有助于理解合作,但适用的情况有限,利他惩罚日益被接受为合作进化的机制,当没有利他惩罚机制时,则不会有合作的进化,加入社会结构和利他惩罚时,合作很容易发展,无标度网络结果表明:节点度分布决定合作是否会在结构化群体中进化方面起着重要作用。
Tomasello et al.(2012) 认为人类合作形式都源于互利,人类合作有两个关键步骤:第一,强制协同觅食。猎鹿博弈说明个人必须与他人合作才能收益。第二,合作技能和动机被扩大到一般的群体生活。随着人类变得相互依赖,人类的认知和社会性变得越来越合作和利他。Sterelny (2014) 认为人类合作经历两次转变,第一阶段是从类人猿的社会生活过度到更新世的合作觅食生活 (利益驱使),第二阶段是从全新世早期向复杂等级社会转变的社会契约稳定性 (the stability of the social contract)(群体冲突所驱动的群体选择)。
社会偏好是利他合作的直接原因。韦倩 (2012) 认为社会得以存在的原因就是行为人的亲社会情感,梳理文献发现在神经科学和行为实验中找到亲社会偏好存在的证据:当亲社会行为发生时,可以观察控制激励的中脑区域的纹状体显著激活;最后通牒博弈、独裁者博弈、公共物品博弈可以说明亲社会偏好的存在。从进化的视角看基因 - 文化协同演化、社会认同和社会学习效应可以解释亲社会偏好何以存在。
亲社会行为与大脑区域有关,特别是前额叶皮层、前脑岛、前扣带回皮层和杏仁核,与亲社会行为相关或因果关系,Luo (2018) 根据神经科学的发现,描述了亲社会行为的共同神经回路。Henrich and Boyd (2001) 证明亲社会基因在文化群体选择改变的环境中广泛传播。Benabou and Tirole (2006) 发展了亲社会行为理论,认为行为主体的亲社会或反社会行为反映了三种动机的内在混合:内在动机、外在动机和声誉,这些行为动机能从其选择和背景中进行推断。
Bell (2010) 认为是文化而不是基因促进了大规模人类亲社会性的进化。Francois etal (2018) 从工作场所和实验室中支持了文化群体选择是人类亲社会的贡献者,企业间的竞争使合作行为增加,实验证明了跨群体竞争可以改变受试者行为和表面上的信念,研究最终表明,工作场所是合作亲社会行为的重要渠道。李诗田、宋献中 (2015) 认为管理者具有亲社会偏好特征的企业,会承担更多的社会责任。
群体选择理论可以解释群体层面的亲社会行为。行为模式随着物种的进化而进化,在生物进化中,合作群体比不合作群体更容易被选择,但群体选择的条件是否在生物进化中得到满足,生物学家一直存在争论,例如约翰・梅纳德・史密斯 (2010) 认为群体选择所需条件苛刻,发生群体选择的可能性极低。West ct al.(007) 认为亲缘选择与群体选择只是计算进化过程或将其概念化的不同方式,实践中亲缘选择更适用,而群体选择经常产生负面后果,群体选择应该是一种潜在有用的、非正式的、概念化的问题解决方式,不是一种一般的进化方式。
虽然关于群体选择在生物学中存在争议,但最近以来特别是在文化演化领域得到了广泛的支持。索伯与威尔逊 (Sober and Wilson,1998) 通过构建数学模型证明利他主义进化的合理性。但张涛 (2012) 认为索伯和威尔逊的数学模型过分注重数学符号,脱离了现实世界,并在此基础上,提出利他行为进化模型,利他者付出给群体带来更高回报时,拥有利他群体会有进化优势,这种进化优势与利他者在群体的比率正相关,利他者本身所持有防御机制使进化优势得到保持。
王健、彭晓娟 (2008) 通过构建博弈模型证明利他主义者在总体中的频率上升。亲缘选择理论、进化博弈论等群体选择的替代理论,可以和群体选择相容,Hamilon and Humilton (1975) 认为亲缘选择是群体选择的一个特例,重复的群体选择可以增加群体中的亲缘关系。Boyd and Richerson (2009) 认为快速的文化适应导致群体之间存在差异,群体竞争导致有竞争能力的行为传播,群体内的自然选择有利于产生新的、更亲社会动机的基因,社会制度增加了这种基因的延续。
赵妍妍 (2011) 分析认为群体选择可以完整地解释生物利他主义,但群体选择并没有穷尽对生物利他现象的解释。Rachlin (2019) 认为行为进化 (以及文化进化) 满足了群体选择的条件,利他主义可以通过行为进化的群体选择学习。Wang et al.(2021) 提出了群体间策略选择的进化博弈模型,研究了群体的选择偏好、群体规模、乘数因子、模仿强度和群体间竞争对群体协同进化的影响,结果表明,无偏好的群体策略的群体间选择对合作进化有积极的影响,带有偏好的策略选择可以有效提高群体的合作效率。黄凯南 (2008) 认为群体选择解释了制度演化速度。
文化 - 基因协同进化解释了不同文化中的合作行为。人类从动物世界脱颖而出,关键在于人类的语言能力和理性能力的获得,由此产生了经验代际累积进化和文化的出现。语言能力、道德规则为个体间高效率信息交流和个体间结构化准备了条件,从而使一种全新 “群体”—— 文化群体的出现变得可能。从早期人类的许多部落到今天众多的民族国家实质上都是不同的文化群体。
文化的产生加速了人类进化,人类文化进化建立在生物进化基础上,哈耶克 (2000) 认为习俗、道德、法律和其他文化都受到群体选择的影响,如果它们携带有利于人类群体生存和扩大的因素,通常会存活和被复制。文化选择在解释人类合作中具有重要作用,Henrich (2004) 认为已有的基因进化方法不能解释人类的亲社会行为,提出基因 - 文化共同进化 —— 基于文化和基因传播之间相互作用的亲社会进化方法,文化进化过程为自然选择提供了条件,从而有利于亲社会基因,人类具有学习能力 (保证高保真模仿),而这些能力会导致基因 - 文化共同进化。
基因 — 文化共同进化通过文化传播的进化和差异人口增长之间相互作用 (Richerson and Boyd,2005)。虽然 Smith (2020) 综述现有研究认为文化群体选择没有为人类合作提供一个满意的解释,但文化群体选择是不可否认的事实。文化过程可以重塑个体面临的选择压力,从而有利于先前没有优势的行为特征的进化 (Efferson et al.,2008)。Richerson et al.(2010) 研究表明文化进化在基因进化中起着积极的主导作用,文化通常比基因进化得更快,文化创造了环境,使基因面临新的选择,许多人的基因在当下正发生着变化。
文化 — 基因协同演化被认为 “最有望弥合社会科学与生物学的理论”(约瑟夫・亨里奇,2018)。Handley and Mathew (2020) 的实验表明,社会中的合作规范是群体选择在文化差异的影响下演变而来的。文化对基因的影响可以表示为:文化→精神发育的表观遗传规则→基因 (拉姆斯登、威尔逊,2016;Henrich,2016;史少博,2009)。
学者从不同的角度分析人类合作何以可能,但并无定论。目前群体选择仍然是主流的观点,批判群体选择观点主要认为其犯了 “平均主义谬误”。本文在 Sober and Wilson (1998) 构建的数学模型的基础上,以普莱斯方程解释群体选择理论,当存在多个群体的竞争和选择时,利他主义可以进化。进一步拓展进行多次迭代,基于普莱斯方程的数值模拟和演化仿真,说明群体选择解释利他主义进化的合理性,以及进化中存在的规律。并将基因演化拓展到文化演化,在以上研究的基础上,我们认为利己、利他概念具有误导性,因为犯了 “直观 - 近因谬误”,提出将 “合作行为” 替换 “利他主义行为”,“非合作行为” 替换 “利己主义行为”。
二、从辛普森悖论 (Simpson’s Paradox) 到群体选择
功利主义集大成者边沁 (Bentham) 认为,大自然使人类服从两个至高的主宰:痛苦和快乐。日常消费品的享乐主义不兼容利他行为;心理上的享乐主义扩展到带来心理满足的利他行为。
从个体选择上的适应度 (fitness) 视角来看,利他主义行为是非适应的:斑马跑得快,更可能逃避天敌的追杀,因此具有更好的生存能力,统计上看拥有更多后代。自然选择倾向于激励自利,扼杀利他行为。与适应性进化紧密联系的概念 —— 适应器 (adaptation):自然选择塑造了适应器,即选择单位的性状。个体适应器表现为奔跑能力等。人类 (个体) 的适应器 = 动物意义上的全部适应器 (例如自利本能)+ 人类特有的文化本能、情感系统、心理倾向、理性能力、对规则的遵从。如果将其他适应器视为给定背景,那么我们可以用追求目标 (purpose - seking) 和规则遵循 (rule - following) 两个特征来刻画人的行为。
人们对自然选择应用在一个层级还是多个层级存在广泛争议。自然选择机制应用在个体层级表现为个体选择;自然选择机制应用在群体层级表现为群体选择。一般认为,个体选择中自利行为被自然选择青睐;群体选择中,导致群体适应度优势的个体性状被自然选择青睐,这种性状导致个体间结构化有利于群体合作剩余的产生,而群体合作剩余平均到个体时,个体收益大于采取自利行为的收益。人类文化群体 (Human Cultural Group) 与群体选择。人类的诞生 —— 心智革命 —— 文化演化介入和叠加到原来的动物生物进化 (基因进化) 中来,足够且必须单独处理,这就是文化演化,或者说 “人类文化群体” 演化。当我们谈论群体选择时,总是考虑有心智能力、理性、文化继承和文化传统的人类,人类总是生活在不同层级嵌套的、大大小小的群体中,最底层的群体就是 “家庭”,最大的群体是跨国家联盟,例如欧盟。
达尔文 (1983) 在《人类的由来》中提出一个初步但明确的 “群体选择” 模型,一些部落 (讲道德的个体充斥) 胜过另一些部落 (相对不那么讲道德的个体充斥)。达尔文刻画 “道德” 使用的概念:爱国精神、忠诚、服从、勇敢、同情心、乐于助人,为共同利益牺牲自……。达尔文明确地意识到自然选择机制可能操作在 “人类文化群体” 中,并提出同情心 —— 社会性本能中最重要的因素,当初无疑通过自然选择而发展起来的。不过达尔文写作此书的目的旨在讨论人与其他动物在生物演化上的连续性,所以他的群体选择思想中的自然选择既应用在生物学遗传单位上,又应用在部落这样的文化群体单位上。讨论自然选择可能应用在文化群体上时,达尔文特地使用了 Natural Selecion as Affecting Civilised Nations 小标题,以示强调。随后正文中达尔文使用了 the agency of natural selection on civilised nations 这样的明确表达 (达尔文,1983:167)。
群体选择的理论假设,当存在多个群体和群体间竞争,帮助自己群体内成员的行为,得到进化 (被群体选择机制 “选择” 出来),因为除了个体之间的竞争以外,存在群体之间的竞争。奔跑的斑马群不是 “群体”,但一个人类部落,一个企业,一个民族国家是真实 “群体”(存在个体间紧密联系和分工结构)。
(一)辛普森悖论
为了对群体选择理论有一个直观的了解,我们看看统计学著名的 “辛普森悖论”。20 世纪 70 年代加州大学伯克利分校研究生招生中,各院系女性录取率都大于或等于男性;但总体上,女性录取率低于男性。我们通过表 3 数值模拟来再现大学招生版 “辛普森悖论”。
| 经济学院 | 文学院 | |
|---|---|---|
| 申请人数 | 90 女,10 男 | 10 女,90 男 |
| 录取率 | 0.30 | 0.60 |
| 录取男性 | 3 | 54 |
| 录取女性 | 27 | 6 |
总体规模 (经济学院 + 文学院)
| 申请总人数 | 100 + 100 = 200 |
|---|---|
| 男性录取率 | (3 + 54) / 200 = 0.29 |
| 女性录取率 | (27 + 6) / 200 = 0.165 |
为什么会出现 “辛普森悖论”?Sober 和 Wilson (1998) 指出,这里针对个体层级的频率计算,隐藏了一个完全忽略 “群体、群体性状、群体适应度” 而导致 “合作剩余” 被放大的 “平均主义谬误”(the averaging fallacy)。
通过辛普森悖论直观呈现的群体选择的 “魔法” 表明,当存在多个群体,且群体之间存在有限交流和竞争(即相对群体隔离),从而存在群体选择时(下次选择不是个体完全混合随机重组新一轮群体,而是存在相对稳定的群体隔离),利他主义行为得到进化。
(二)利他主义如何可能得到进化?
1.利他主义在自然选择中倾向于灭绝:单群体模型
个体适应度包括生存能力和繁殖能力,在这个模型中,利他行为只影响繁殖,所以后代数量是衡量适应度的标准 (Sober 和 Wilson,1998)。
假设:包含 n 个个体的群体,群体中只有两种基因编码的性状:A-- 利他;S-- 利己。利他者频率 P ,利己者频率 1 − p 1 - p 1−p 。这样群体中的利他主义者个数 n p np np,利己主主义者个数 n ( 1 − p ) n (1 - p) n(1−p) 。
平均适应度:不存在利他主义者时,所有个体拥有相同数量后代,平均适应度为 X。
突变出利他主义者,利他主义行为会减少 c 个自己的后代,并为群体中利益接受者增加 b 个后代。
那么,利己主义者的适应度 W s W_{s} Ws,利他主义者的适应度 W a W_{a} Wa 用如下方程式表示:
W a = X − c + [ b ( n p − 1 ) n − 1 ] ( 1 ) W_{a}=X - c+\left [\frac {b (np - 1)}{n - 1}\right] (1) Wa=X−c+[n−1b(np−1)](1)
W s = X + [ b n p n − 1 ] ( 2 ) W_{s}=X+\left [\frac {bnp}{n - 1}\right] (2) Ws=X+[n−1bnp](2)
每一个利他主义者都付出其行为的代价 ( − c ) (-c) (−c),但同时也是群体中其他利他主义者 ( n p − 1 ) (np - 1) (np−1) 利益的可能接受者。由于其他利他主义者在 ( n − 1 ) (n - 1) (n−1) 个个体中分配他们的利益,每个利他主义者经历的总期望利益为 b ( n p − 1 ) n − 1 \frac {b (np - 1)}{n - 1} n−1b(np−1) 。非利他主义者不经历利他主义的代价,同时也可能成为所有 n p np np 利他主义者的接受者。很明显, W a W_{a} Wa 总是小于 W s W_{s} Ws,所以利他主义者总是在这个种群中被选择 (Sober 和 Wilson,1998)。
参数赋值,数值模拟上述方程, n = 100 n = 100 n=100, p = 0.5 p = 0.5 p=0.5, X = 10 X = 10 X=10, b = 5 b = 5 b=5, c = 1 c = 1 c=1,代入上述方程计算得:
W a = 10 − 1 + 5 ∗ 49 99 = 11.47 W_{a}=10 - 1+5 * \frac {49}{99}=11.47 Wa=10−1+5∗9949=11.47
W s = 10 + 5 ∗ 50 99 = 12.53 W_{s}=10+5 * \frac {50}{99}=12.53 Ws=10+5∗9950=12.53
后代总数: n ′ = n [ p W a + ( 1 − p ) W a ] = 100 ∗ [ 0.5 ∗ 11.47 + 0.5 ∗ 12.53 ] = 1200 n'=n [pW_{a}+(1 - p) W_{a}]=100 *[0.5 * 11.47+0.5 * 12.53]=1200 n′=n[pWa+(1−p)Wa]=100∗[0.5∗11.47+0.5∗12.53]=1200
后代中利他主义者频率 p ′ = n p W a n ′ = 100 ∗ 0.5 ∗ 11.47 1200 = 0.47791667 p'=\frac {npW_{a}}{n'}=100 * 0.5 * \frac {11.47}{1200}=0.47791667 p′=n′npWa=100∗0.5∗120011.47=0.47791667
如果假定:两种类型行为者死亡率相同,且群体规模限制在 100,那么第二代中利他主义者个数约为 48,利己主义者个数约为 52。
结论是:一个群体情形,迭代次数足够多时,利他主义者频率趋势为 (0.5,0.48,……)→0,数量趋势为 (50,48,…)。
西蒙 (2009:43) 提出一个更简洁模型说明利他主义者为何在演化中被清除。
假设: X 一行为者的与生俱来的适应度,或者平均适应度;
E s Es Es 一 自利者适应度;
F a Fa Fa 一 利他者适应度;
c 一 利他行为付出的成本;
b 一 是利他行为量,b 是 c 的函数, b ( c ) b (c) b(c) ;
p 一 利他行为者的频率;
那么:利主义者和和利他主义者的适应度分别是
s = X + b ( c ) p s=X + b (c) p \quad s=X+b(c)p (3)
a = X − c + b ( c ) p a=X - c + b (c) p a=X−c+b(c)p (4)
在 (3) 与 (4) 式中,利他主义者每一代的适应度都比利已主义者低,长期中必然倾向于被清除。
2.考虑多群体情形:一个奇怪的结论
一旦我们考虑多群体,并把 “群体选择” 引入到演化进程,情况就可能出现反转。我们考虑两群体,按照上述单群体模型中设定的适应度,对参数赋值进行数值模拟,计算选择前后的群体规模和比率,我们就会惊奇地发现,利他主义者在种群层面的比例比选择前的初始频率提升了。不过,在文化演化领域中的群体选择,“利他主义” 的含义需要修正,它实际上是指基于专业化、分工和交换的 “合作行为”(见后文第六部分的专门讨论)。表 4 是根据 Sober 和 Wilson (1998) 群体选择模型演变而来的数值模拟,对群体的初始规模分别赋值 100000,10000,以看出合作者频率是上升的。
表 4
2 群体 2 层级群体选择模型数值模拟
| 参数 | 群体 1 | 群体 2 | 说明 |
|---|---|---|---|
| 选择前群体规模 ( n ) (n) (n) | 100000 | 100000 | |
| 合作者的频率 ( p ) (p) (p) | 0.20 | 0.80 | |
| 平均适应度 X | 4 | 4 | |
| 受惠者从合作者那里获得收益 b | 3 | 3 | |
| 合作者合作行为的成本 c | 1 | 1 | |
| 合作者适应度 | 4 − 1 + 3 ∗ ( 100000 ∗ 0.2 − 1 ) / ( 100000 − 1 ) = 3.60 4 - 1+3*(100000*0.2 - 1)/(100000 - 1)=3.60 4−1+3∗(100000∗0.2−1)/(100000−1)=3.60 | 4 − 1 + 3 ∗ ( 100000 ∗ 0.8 − 1 ) / ( 100000 − 1 ) = 5.40 4 - 1+3*(100000*0.8 - 1)/(100000 - 1)=5.40 4−1+3∗(100000∗0.8−1)/(100000−1)=5.40 | |
| 自我中心主义非合作者的适应度 W D W_{D} WD | 4 + 3 ∗ 100000 ∗ 0.2 / ( 100000 − 1 ) = 4.60 4+3*100000*0.2/(100000 - 1)=4.60 4+3∗100000∗0.2/(100000−1)=4.60 | 4 + 3 ∗ 100000 ∗ 0.8 / ( 100000 − 1 ) = 6.40 4+3*100000*0.8/(100000 - 1)=6.40 4+3∗100000∗0.8/(100000−1)=6.40 | |
| 选择后群体规模 ( n ′ ) (n') (n′) | 20000 ∗ 3.6000 + 80000 ∗ 4.6000 = 440000 20000*3.6000 + 80000*4.6000 = 440000 20000∗3.6000+80000∗4.6000=440000 | 80000 ∗ 5.4000 + 20000 ∗ 6.4000 = 560000 80000*5.4000 + 20000*6.4000 = 560000 80000∗5.4000+20000∗6.4000=560000 | |
| 选择后合作者的频率 | 100000 ∗ 0.2 ∗ 3.6000 / 440000 = 0.16 100000*0.2*3.6000/440000 = 0.16 100000∗0.2∗3.6000/440000=0.16 | 100000 ∗ 0.8 ∗ 5.4000 / 560000 = 0.77 100000*0.8*5.4000/560000 = 0.77 100000∗0.8∗5.4000/560000=0.77 | 合作者频率下降 |
总体规模 (群体 1 + 群体 2)
| N N N | 100000 + 100000 = 200000 100000 + 100000 = 200000 100000+100000=200000 | |
|---|---|---|
| p 1 + 2 p_{1 + 2} p1+2 | ( 0.2 ∗ 100000 + 0.8 ∗ 100000 ) / 200000 = 0.50 (0.2*100000 + 0.8*100000)/200000 = 0.50 (0.2∗100000+0.8∗100000)/200000=0.50 | |
| N r N^{r} Nr | 440000 + 560000 = 1000000 440000 + 560000 = 1000000 440000+560000=1000000 | |
| p 1 + 2 ′ p_{1 + 2}' p1+2′ | ( 0.1637 ∗ 440000 + 0.7714 ∗ 560000 ) / 1000000 = 0.504 (0.1637*440000 + 0.7714*560000)/1000000 = 0.504 (0.1637∗440000+0.7714∗560000)/1000000=0.504 | 合作者频率上升 |
三、不借助群体选择来解释利他主义行为:亲缘选择与汉密尔顿法则
在动物世界和人类世界,生物学家倾向于认为,用亲缘选择理论解释了任何利他主义行为。然而,在人类世界,一再观察到和人们感知到的真利他主义行为发生在非亲缘关系的人际间,因而提出,亲缘选择理论至少不能解释全部利他主义行为。
(一)汉密尔顿法则
首先我们看亲缘选择理论的数学表示,这个数学不等式由汉密尔顿提出,因此通常被称为 “汉密尔顿法则”。
假设:
r 一 个体 (等位基因间) 亲缘关系系数;
b 一 个体 (等位基因) 行动的收益;
c 一 个体 (等位基因) 行动的成本;
那么:满足 r b > c rb>c rb>c 时,利他主义行为得到进化。
汉密尔顿法则可以变换为,当满足 r > c b r>\frac {c}{b} r>bc 或者 r < b c r<\frac {b}{c} r<cb 时,利他主义得到进化。与新古典经济学的行为假设相联系。
当 r = 1 r = 1 r=1 时,行为者就是自己。此时,当 1 ∗ b c 1 * \frac {b}{c} 1∗cb 即 b c \frac {b}{c} cb 时,个体采取行动。
当 r = 0.5 r = 0.5 r=0.5 时,当 b > 2 c b>2c b>2c 时,个体采取行动,在生物学意义上,就是父母对子女的利他主义行为的条件 —— 收益大于成本的 2 倍,父母帮助孩子。
当 r → 0 r \to 0 r→0 时,就是匿名的陌生人世界,此时 b 需要无穷大,个体才会采取帮助行为。但是,回到人类社会现实,在匿名陌生人世界,当考虑人类心理倾向 (具有近似本能的性质,无论这种准本能倾向是来自于基因遗传还是后天文化习得),道德约束、社会规范的伦理约束,以及宗教信仰等因素时,r 不会趋近于 0,这样,相应的个体实施利他主义行为的 b 远大于 2 c 2c 2c,但不会无穷大。
上述讨论我们可以得出如下结论:新古典经济学的 “经济人假设” 是汉密尔顿方程的特殊情形。汉密尔顿方程对利他主义进化的必要条件的说明是不充分的。
(二)对汉密尔顿方程的改造:用 “交往系数” 替代 “亲缘关系系数”
从人类早期的小规模社会进化到 “大社会” 也就是哈耶克著名的 “扩展秩序” 以后,人类分工合作的纽带显然大大超越了亲属和族群,用亲缘选择来解释大规模分工合作显然不符合现实,我们用交换系数来替换亲缘关系系数 (Axelrod 和 Hamilton,1981;Lehmann 和 Keller,2006;Boyd,2017):
假设: T 为合作者之间平均交往次数;
b 为受益者在群体中的适应度增加;
c 为利他行为的成本。
那么,改造后的汉密尔顿方程为:
( 1 − 1 T ) b > c (1-\frac {1}{T}) b>c (1−T1)b>c,或者,用收益成本比来表示为 (T 无穷大时等于自己;T 非常大时为近亲属的情形;T 为 1 时为陌生人):
T T − 1 < b c \frac {T}{T - 1}<\frac {b}{c} T−1T<cb
我们取两个简单的模拟值,例如交往次数为 2 和 100,那么利他合作得以维持的条件是:
2 2 − 1 < b c \frac {2}{2 - 1}<\frac {b}{c} 2−12<cb,即 2 < b c 2<\frac {b}{c} 2<cb (当交往稀疏时,维持合作的收益成本比大于 2)。
100 99 < b c \frac {100}{99}<\frac {b}{c} 99100<cb,即 1.0101 < b c 1.0101<\frac {b}{c} 1.0101<cb (当交往非常密时,维持合作的收益成本比要求较低)。
亲属选择和互惠利他主义为小群体持续合作提供了解释,但这些观点不足以理解大规模社会中亲社会行为和大范围社会合作秩序。用交往系数替代亲缘关系系数是试图在亲缘选择理论框架下为超越亲缘关系的大范围合作秩序提供解释,但这种借助于 (局限于) 基因联系的群体层级分析并不成功。
四、解决 “群体选择” 的首个数学模型:普莱斯方程 (Price Equation)
普莱斯方程形成了亲缘选择和群体选择的基础 (Lehtonen,2020)。普莱斯方程把群体内的个体选择与群体间的群体选择整合在一个方程中。一方面,在群体内的选择中,合作者个体的个体适应度降低,但所在的群体的规模会扩张。另一方面,群体间存在群体选择,那些扩张的群体相对那些不能扩张甚至缩小的群体占优势,在种群层面,合作者的频率会上升。普莱斯方程表达的合作者的频率是两项的算术和:第一项是单个群体内的频率平均变化;第二项是群体适应度差异引起的频率平均变化。这样,合作者的净适应度是,群体内个体选择导致的个体适应度下降与群体间群体选择导致的个体适应度上升的最终净效果。普莱斯方程清晰地将群体内的选择力量与群体间选择力量区分开来,然后加起来得到个体适应度的净效果。
假设:
m : 种群有 m 个群体,群体间存在严格边界但有限度的相互交流 (例如移民)
选择前:
n i n_{i} ni : 每个群体 i 的初始规模;
p i p_{i} pi : 合作者的初始基因频率;
n : 表示平均群体规模;
p : 表示平均合作者频率。
选择后:
n i ′ n'_{i} ni′ : 选择后群体规模;
p i ′ p'_{i} pi′ : 选择后合作者的频率;
n ′ n' n′ : 表示平均群体规模;
p ′ p' p′ : 表示平均合作者频率;
群体收益 s i = n i ′ / n i s_{i}=n_{i}' /n_{i} si=ni′/ni
因为这里考虑的是合作者基因,所以 n i ′ > n i n_{i}'>n_{i} ni′>ni ; p i ′ < p i p'_{i}<p_{i} pi′<pi
普莱斯方程:
1.种群层次
选择前合作者基因频率为:
= ( ∑ n i p i ) / ( ∑ n i ) = p + c o v ( n , p ) / n =\left (\sum n_{i} p_{i}\right)/\left (\sum n_{i}\right)=p + cov (n, p)/n =(∑nipi)/(∑ni)=p+cov(n,p)/n (5)
选择后合作者基因频率为:
P ′ = ∑ n i ′ p i ′ ∑ n i ′ = p ′ + c o v ( n ′ , p ′ ) / n ′ P'=\frac {\sum n_{i}'p_{i}'}{\sum n'_{i}}=p'+cov\left (n', p'\right)/n' P′=∑ni′∑ni′pi′=p′+cov(n′,p′)/n′ (6)
2.种群层级合作者频率变化
选择后,合作者基因频率变化定义为 Δ P = P ′ − P \Delta P=P' - P ΔP=P′−P,
Δ P = a v e n ′ ( Δ p ) + c o v n ( s , p ) a v e n s \Delta P=ave_{n'}(\Delta p)+\frac {cov_{n}(s, p)}{ave_{n} s} ΔP=aven′(Δp)+avenscovn(s,p) (7)
第一项是平均群体中基因频率的改变,由选择后的群体规模 ( n ′ ) (n') (n′) 加权计算,是衡量群体内的选择情况。第二项包含了群体收益 ( s ) (s) (s) 和选择前 (当群体规模为 n) 群体内利他主义者出现频次 ( p ) (p) (p) 之间的协方差,是衡量群体间的选择情况 (Sober 和 Wilson,1998:74)。
3.群体选择模型的一般形式
我们可以通过一般化模型说明合作者在种群中扩张的条件:假设种群中有 m m m 个群体,每个群体有 n n n 个个体,分别由合作者( C C C)和自我中心主义的非合作者( D D D)组成,第 i i i 组合作者频率为 P i P_i Pi,合作者适应度: W c i = X − c + [ b ( n p i − 1 ) n − 1 ] W_{ci} = X - c + \left [\frac {b (np_i - 1)}{n - 1}\right] Wci=X−c+[n−1b(npi−1)],自我中心主义非合作者的适应度: W D i = X + b n p i n − 1 W_{Di} = X + \frac {bnp_i}{n - 1} WDi=X+n−1bnpi,选择后的群体规模 n i ′ = n [ p i W c i + ( 1 − p i ) W D i ] = n X + p i n ( b − c ) n'_i = n [p_iW_{ci} + (1 - p_i) W_{Di}] = nX + p_in (b - c) ni′=n[piWci+(1−pi)WDi]=nX+pin(b−c),选择后合作者的频率 p i ′ = n p i W c i n i ′ p'_i = \frac {np_iW_{ci}}{n'_i} pi′=ni′npiWci。可以证明(由于 ( 1 − p i ) ( b n − 1 + c ) > 0 (1 - p_i)(\frac {b}{n - 1} + c) > 0 (1−pi)(n−1b+c)>0,所以 W c i W c i + ( 1 − p i ) ( b n − 1 + c ) < 1 \frac {W_{ci}}{W_{ci} + (1 - p_i)(\frac {b}{n - 1} + c)} < 1 Wci+(1−pi)(n−1b+c)Wci<1, p i W c i W c i + ( 1 − p i ) ( b n − 1 + c ) < p i \frac {p_iW_{ci}}{W_{ci} + (1 - p_i)(\frac {b}{n - 1} + c)} < p_i Wci+(1−pi)(n−1b+c)piWci<pi):
p i ′ = n p i W c i n i ′ = n p i W c i n [ p i W c i + ( 1 − p i ) W D i ] = p i W c i [ p i W c i + ( 1 − p i ) W D i ] = p i W c i W c i + ( 1 − p i ) ( b n − 1 + c ) < p i ( 8 ) p'_i = \frac {np_iW_{ci}}{n'_i} = \frac {np_iW_{ci}}{n [p_iW_{ci} + (1 - p_i) W_{Di}]} = \frac {p_iW_{ci}}{[p_iW_{ci} + (1 - p_i) W_{Di}]} = \frac {p_iW_{ci}}{W_{ci} + (1 - p_i)(\frac {b}{n - 1} + c)} < p_i \quad (8) pi′=ni′npiWci=n[piWci+(1−pi)WDi]npiWci=[piWci+(1−pi)WDi]piWci=Wci+(1−pi)(n−1b+c)piWci<pi(8)
可见,合作者频率在群体内是逐代下降的。
而在种群层级,种群中合作者频率扩张的一般条件:
X i X_{i} Xi 表示第 i i i 组合作者的个数, X i = n p i X_{i}=np_{i} Xi=npi
E ( X ) E (X) E(X) 为群体中合作者期望数量;
V a r ( X ) Var (X) Var(X) 为群体间的合作者数量的方差; E ( X ) ( n − E ( x ) ) E (X)(n - E (x)) E(X)(n−E(x)) 为群体内两种类型个体数量的方差。
选择前合作者的频率 p = E ( X ) n p=\frac {E (X)}{n} p=nE(X)
选择后合作者频率 p ′ = ∑ i = 1 m p i ′ n i ′ ∑ i = 1 m n i ′ = ∑ i = 1 m p i W c i ∑ i = 1 m [ p i W c i + ( 1 − p i ) W D i ] p'=\frac {\sum_{i = 1}^{m} p_{i}'n_{i}'}{\sum_{i = 1}^{m} n_{i}'}=\frac {\sum_{i = 1}^{m} p_{i} W_{ci}}{\sum_{i = 1}^{m}[p_{i} W_{ci}+(1 - p_{i}) W_{Di}]} p′=∑i=1mni′∑i=1mpi′ni′=∑i=1m[piWci+(1−pi)WDi]∑i=1mpiWci ,计算可得选择后合作者频率增加 ( p ′ > p ) (p'>p) (p′>p) 的条件为:
( n − 1 ) ( 1 − p ) ∑ i = 1 m p i n ∑ i = 1 m p i 2 − ∑ i = 1 m p i − ( n − 1 ) p ∑ i = 1 m p i < b c \frac {(n - 1)(1 - p)\sum_{i = 1}^{m} p_{i}}{n\sum_{i = 1}^{m} p_{i}^{2}-\sum_{i = 1}^{m} p_{i}-(n - 1) p\sum_{i = 1}^{m} p_{i}}<\frac {b}{c} n∑i=1mpi2−∑i=1mpi−(n−1)p∑i=1mpi(n−1)(1−p)∑i=1mpi<cb (9)
进一步考察式 (9),因为 ∑ i = 1 m p i = m E ( X ) n \sum_{i = 1}^{m} p_{i}=\frac {mE (X)}{n} ∑i=1mpi=nmE(X) , ∑ i = 1 m p i 2 = m E ( X 2 ) n 2 \sum_{i = 1}^{m} p_{i}^{2}=\frac {mE (X^{2})}{n^{2}} ∑i=1mpi2=n2mE(X2) ,所以,式 (9) 可以表达为:
( n − 1 ) E ( X ) ( n − E ( X ) ) n V a r ( X ) − n E ( X ) + E 2 ( X ) < b c \frac {(n - 1) E (X)(n - E (X))}{nVar (X)-nE (X)+E^{2}(X)}<\frac {b}{c} nVar(X)−nE(X)+E2(X)(n−1)E(X)(n−E(X))<cb (10)
由式 (10) 可知,收益成本比与合作者个数的群间方差 V a r ( X ) Var (X) Var(X) 以及群内方差 E ( X ) ( n − E ( X ) ) E (X)(n - E (X)) E(X)(n−E(X)) 相关,群间合作者个数占比差异较大,群内合作者个体差异较小,即群体间的适应度差异 (偏爱合作性行为的力量) 必须强到超过群体内诸个体间的适应度差异 (偏爱非合作性行为的力量),则合作者行为就越有可能在种群中占有利位置,这是合作者在多群体模型中得到进化的必要条件 (Sober 和 Wilson,1998:26)。出现这种现象的原因是,随着群体扩大,合作者行为会提高群体合作收益,也就是说,在群体选择中,合作性行为具有适应性,群体选择会青睐增加群体适应度的行为。
4.群体选择的演化仿真
本文运用 java 的 Eclipse 开发工具计算群体中合作者频率迭代变化,假设种群包含两个具有相同规模的群体,群体 1 和群体 2, P 1 P_{1} P1 表示群体 1 迭代前的合作者频率, P 2 P_{2} P2 表示群体 2 迭代前的合作者频率。由于群体 1 与群体 2 是两个完全相同的群体,故设 P 1 < P 2 P_{1}<P_{2} P1<P2 。所以可设:
P 1 + P 2 2 < P 1 ′ n 1 ′ + P 2 ′ n 2 ′ n 1 + n 2 ′ \frac {P_{1}+P_{2}}{2}<\frac {P_{1}'n_{1}'+P_{2}'n_{2}'}{n_{1}+n_{2}'} 2P1+P2<n1+n2′P1′n1′+P2′n2′ (11)
不等式左边的 P 1 P_{1} P1、 P 2 P_{2} P2 是迭代前频率的初始值,右边的 P 1 ′ P_{1}' P1′、 P 2 ′ P_{2}' P2′ 是迭代后的频率值,同样, n 1 ′ n_{1}' n1′、 n 1 ′ n_{1}' n1′ 是迭代后群体规模值。
所有的群体预设了相同的基准适应度,成本和收益在这个基准适应度的基础上进行加减。最终进化的产物取决于反对合作者的群体内选择和支持合作者群体间选择共同产生的相对强度。本实验设置平均适应度 X X X 为 2 个单位,合作者行动时成本 c c c 为 1 个单位, b b b 比 c c c 大 10 倍在现实生活中很少见,但当个人行为攸关群体生死存亡的时候,此时,利他行为做出行动,会使其他成员后代增加超过 c c c 的 10 倍, b b b 值可以取无限大,考虑可操作性,设置 b b b 不超过 1000。观察种群中合作者频率上升时群体中利他收益 b b b 值变化以及两个群体中初始合作者频率 ( p 1 p_{1} p1 与 p 2 p_{2} p2 ) 变化情况,得到几点结论:
结论 1: p 1 p_{1} p1 固定时, p 2 p_{2} p2 与 p 1 p_{1} p1 的差距越大,如果满足迭代后种群中合作者频率上升,则对 b b b 的要求越小。
结论 2:随着迭代次数的增加, p 1 p_{1} p1 和 p 2 p_{2} p2 相同的情况,则需要更大的 b b b 值的支持,种群中合作者才能得到进化 。
结论 3:相同条件下,初始群体规模对 P P P 和 b b b 最小取值影响不大,随着迭代次数或者合作者频率的增加,初始规模的影响小于群体内部机制的影响,当迭代次数很少或者群体内合作者频率很小,初始规模的影响才显现,此时初始规模越大,则需要的 b b b 减小。
结论 4:当合作者做出行动时,成本 c c c 增加,则所需最小 b b b 值也成倍增加。
结论 5:相同条件下,当合作者做出行动时,平均适应度 X X X 增加,则所需 b b b 值降低,但总体来看,平均适应度 X X X 对 P P P 和 b b b 值的影响不大。
结论 6:考虑固定群体规模与不固定群体规模两种情形,不固定规模相对于固定规模 ( n = 150 n = 150 n=150 , n = 100000 n = 100000 n=100000 ),所需 b b b 值较小。
结论 7:普莱斯方程仅考虑合作成本、接受者收益、群体规模、合作者频率,不涉及真实人类社会中的社会规范内化,遵从、羞愧、愤怒等先天心理机制,以及声誉机制、第三方惩罚等维护合作的各种力量,一旦将这种促进合作的个体和社会的 “执行机制” 考虑进去,即每一次迭代后的合作者规模乘以一个 “加强” 系数 (例如 1.001),则合作者迭代后频率上升所需 b b b 值要求更低。
给定一组初始值,我们观察迭代 50 次后的终值以及演化过程。通过设置群体规模,两个群体的 p p p 初始值,计算满足种群层级合作者频率上升的条件,可以观察到收益成本要求 (表 5)。图 1 是迭代 50 次演化过程数据的图示。
| 参数 | 结果 | 说明 |
|---|---|---|
| 平均适应度 | 1.2 | |
| 利他收益 b b b | 8459 | |
| 合作者行动成本 | 1 | |
| 群体初始规模 n n n | 150 | |
| 两群体中合作者初始频率 p 1 p_{1} p1 和 p 2 p_{2} p2 | p 1 = 0.4 p_{1}=0.4 p1=0.4 p 2 = 0.58 p_{2}=0.58 p2=0.58 | |
| 种群初始规模 N N N | 150 + 150 = 300 150 + 150 = 300 150+150=300 | |
| 种群初始合作者频率 P P P | ( 0.4 ∗ 150 + 0.58 ∗ 150 ) / 300 = 0.49 (0.4*150 + 0.58*150)/300 = 0.49 (0.4∗150+0.58∗150)/300=0.49 | |
| 迭代 50 次后种群中合作者频率 P ′ P' P′ | 0.58 | 合作者频率上升 |
| 迭代 50 次后种群中非合作者频率 P 非 P_{非} P非 | 0.42 | 非合作频率下降 |
将普莱斯方程进一步推广,考虑多群体模式。此时假设种群包含三个群体,群体 1、群体 2、群体 3, P 1 P_{1} P1、 P 2 P_{2} P2、 P 3 P_{3} P3 表示群体 1、群体 2、群体 3 迭代前的合作者频率,群体 1、群体 2 和群体 3 是完全相同的群体,故设 P 1 < P 2 < P 3 P_{1}<P_{2}<P_{3} P1<P2<P3 。
P 1 + P 2 + P 3 3 < P 1 ′ n 1 + P 2 ′ n 1 + P 3 ′ n 3 ′ n 1 + n 2 ′ + n 3 ′ \frac {P_{1}+P_{2}+P_{3}}{3}<\frac {P_{1}'n_{1}+P_{2}'n_{1}+P_{3}'n_{3}'}{n_{1}+n_{2}'+n_{3}'} 3P1+P2+P3<n1+n2′+n3′P1′n1+P2′n1+P3′n3′ (12)
不等式左边的 P 1 P_{1} P1、 P 2 P_{2} P2、 P 3 P_{3} P3 是迭代前的初始值,右边的 P 1 ′ P_{1}' P1′、 P 2 ′ P_{2}' P2′、 P 3 ′ P_{3}' P3′ 是迭代后的合作者频率。同样, n 1 ′ n_{1}' n1′、 n 1 ′ n_{1}' n1′、 n 3 ′ n_{3}' n3′ 是迭代后群体的群体规模。观察种群中合作者频率上升时群体中利他收益 b b b 变化以及三个群体中初始合作者频率 ( P 1 P_{1} P1、 P 2 P_{2} P2、 P 3 P_{3} P3 ) 变化情况。
结论 8:相对于两群体 (合作者频率记为 p 1 p_{1} p1 和 p 2 p_{2} p2 ),当增加一个群体 (此群体合作者频率记为 p 3 p_{3} p3 ) 的 p 3 p_{3} p3 较小时,所需 b b b 值降低,随着增加群体的 p 3 p_{3} p3 增加,所需 b b b 值也增加,当增加的群体的 p 3 p_{3} p3 与两群体中的 p 1 p_{1} p1、 p 2 p_{2} p2 中较大的那个群体相等时,所需最小 b b b 值最大, p 3 p_{3} p3 大于 p 1 p_{1} p1、 P 2 P_{2} P2 时,所需 b b b 值降低 。
表 6 表示给定三个群体的 p p p 初始值,满足种群层级合作者频率上升的条件,收益成本的要求。图 2 是三个群体迭代 50 次演化过程数据的图示。
| 参数 | 结果 | 说明 |
|---|---|---|
| 平均适应度 | 1.2 | |
| 利他收益 b b b | 8459 | |
| 合作者行动成本 | 1 | |
| 群体初始规模 n n n | 150 | |
| 三群体中合作者初始频率 p 1 p_{1} p1、 p 2 p_{2} p2 和 p 3 p_{3} p3 | p 1 = 0.20 p_{1}=0.20 p1=0.20 p 2 = 0.40 p_{2}=0.40 p2=0.40 p 3 = 0.58 p_{3}=0.58 p3=0.58 | |
| 种群初始规模 N N N | 150 + 150 + 150 = 450 150 + 150 + 150 = 450 150+150+150=450 | |
| 种群初始合作者频率 P P P | ( 0.2 ∗ 150 + 0.4 ∗ 150 + 0.58 ∗ 150 ) / 450 = 0.39 (0.2*150 + 0.4*150 + 0.58*150)/450 = 0.39 (0.2∗150+0.4∗150+0.58∗150)/450=0.39 | |
| 迭代 50 次后种群中合作者频率 P ′ P' P′ | 0.58 | 合作者频率上升 |
| 迭代 50 次后种群中非合作者频率 P 非 P_{非} P非 | 0.42 | 非合作者频率下降 |
五、群体选择中的行为讨论
群体选择和多层级选择模型揭示了通往合作秩序的最严格行为条件,即群体中个体牺牲自身适应度 c c c ,提升群中个体其他适应度 b b b ,当且仅当满足式 (10) 时。进一步放松假设,逼近现实,不失一般性,我们假设个体具有 4 种行为,分别为情形 I、情形 II、情形 III,情形 IV,各自满足式 (10) 条件见表 7。
| 行为策略 | c c c 与 b b b | 得到进化的满足式 | |
|---|---|---|---|
| 情形 I | b > 0 b>0 b>0 ; c > 0 c>0 c>0 | 损己利人 | c ( n − 1 ) E ( X ) ( n − E ( X ) ) < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))<b [nVar (X)-nE (X)+E^{2}(X)] c(n−1)E(X)(n−E(X))<b[nVar(X)−nE(X)+E2(X)] |
| 情形 II | b > 0 b>0 b>0 ; c = 0 c=0 c=0 | 不损己利人 | c ( n − 1 ) E ( X ) ( n − E ( X ) ) = 0 < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))=0<b [nVar (X)-nE (X)+E^{2}(X)] c(n−1)E(X)(n−E(X))=0<b[nVar(X)−nE(X)+E2(X)] |
| 情形 III | b > 0 b>0 b>0 ; c < 0 c<0 c<0 | 利己利人 | c ( n − 1 ) E ( X ) ( n − E ( X ) ) < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))<b [nVar (X)-nE (X)+E^{2}(X)] c(n−1)E(X)(n−E(X))<b[nVar(X)−nE(X)+E2(X)] |
| 情形 IV | b < 0 b<0 b<0 ; c < 0 c<0 c<0 | 损人利己 | c ( n − 1 ) E ( X ) ( n − E ( X ) ) < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))<b [nVar (X)-nE (X)+E^{2}(X)] c(n−1)E(X)(n−E(X))<b[nVar(X)−nE(X)+E2(X)] |
情形 I:损己利人。种群中出现损己利人的群体,当损己利人个体做出行动时,个体行为者牺牲适应度为 c c c ,提升群体中其他适应度为 b b b , b > 0 b>0 b>0, c > 0 c>0 c>0 。当 b > 0 b>0 b>0, c > 0 c>0 c>0,且满足式 (13),损己利人可以得到进化:
c ( n − 1 ) E ( X ) ( n − E ( X ) ) < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))<b\left [nVar (X)-nE (X)+E^{2}(X)\right] c(n−1)E(X)(n−E(X))<b[nVar(X)−nE(X)+E2(X)]
根据适者生存的原则,损己利人永远得不到进化,但实际与之相反,从群体选择的角度看,损己利人在群体中具有适应性,可以提高群体利益,群体选择强大的力量使得损己利人得到进化。
情形 II:不损己利人。种群中出现不损己利人的群体,当个体做出行动,个体行为者牺牲适应度为 c c c , c = 0 c = 0 c=0,提升群体中其他适应度为 b b b , b > 0 b>0 b>0 ;当 b > 0 b>0 b>0, c = 0 c = 0 c=0,且满足式 (14),不损己利人可以得到进化:
c ( n − 1 ) E ( X ) ( n − E ( X ) ) = 0 < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))=0<b\left [nVar (X)-nE (X)+E^{2}(X)\right] c(n−1)E(X)(n−E(X))=0<b[nVar(X)−nE(X)+E2(X)]
受惠者可以从不损己利人个体中获得收益,而它们的行为并没有带来损失,进而增加群体的利益,不损己利人可以最大化群体相对适应度的性状而得到进化。
情形 III:利己利人。种群中出现利己利人的群体,当个体做出行动,个体行为者牺牲适应度为 c c c , c < 0 c<0 c<0,提升群体中其他适应度为 b b b , b > 0 b>0 b>0 。当 b > 0 b>0 b>0, c < 0 c<0 c<0,且满足式 (15),利己利人可以得到进化:
c ( n − 1 ) E ( X ) ( n − E ( X ) ) < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))<b\left [nVar (X)-nE (X)+E^{2}(X)\right] c(n−1)E(X)(n−E(X))<b[nVar(X)−nE(X)+E2(X)]
人的本质是利己的或自利的,但同时人格道德和经济道德并存,当 b b b 与 c c c 满足式 (13),群体间适应度差异超过群体内个体间的适应度差异,利己利人群体得到进化。
情形 IV:损人利己。种群中出现损人利己的群体,当损人利己个体做出行动时,个体行为者牺牲适应度为 c c c , c < 0 c<0 c<0,提升群体中其他适应度为 b b b , b < 0 b<0 b<0 。当 b < 0 b<0 b<0, c < 0 c<0 c<0,且满足式 (16),损人利己群体可以得到进化:
c ( n − 1 ) E ( X ) ( n − E ( X ) ) < b [ n V a r ( X ) − n E ( X ) + E 2 ( X ) ] c (n - 1) E (X)(n - E (X))<b\left [nVar (X)-nE (X)+E^{2}(X)\right] c(n−1)E(X)(n−E(X))<b[nVar(X)−nE(X)+E2(X)]
一切生物进化方向都是利己的,“损人” 的目的为了繁殖更多的后代,当满足上述条件,损人利己群体也可以进化。在现实社会中依然存在少数损人利己群体。
值得指出的是,在早期人类社会的经济中与在复杂的现代分工交换经济中,以上四种情形发挥作用空间各不相同,原因是,在部落经济中专业化分工和交换的范围有限,来自于分工结构的合作群体只有数百人的规模,因此很大程度上适用于情形 I,以及相应的道德和社会规范支持;而在复杂的现代分工和交换经济中,市场规模扩大到前所未有的程度,经济秩序主要依赖于 “陌生人之间” 以分工交换实现的合作,因此更大程度上适用情形 II 也就是亚当・斯密阐述过的 “看不见的手” 的情形。但是我们必须指出的是,由于现代人类同时生活在 “三重世界” 之中:(a) 在文化演化进程中;(b) 在观念竞争进程中;© 在公共选择或立法的与个体选择过程中 (Wohlgemuth,2011),因此个体行为的 “净效应” 是以上三重约束下相互加强或抵消的结果。
六、从基因演化到文化演化
(一)群体、性状群体与人类文化群体
广场上的一群人不是 “群体”。群体是具有群体性状的个体集合,因为个体间通过特定的规则而被结构化彼此联系在一起。群体选择现象只存在于人类社会中,因为人类总是以 “群体” 的方式存在,例如部落、民族、民族国家。这些群体都是性状群体,即不同部落、不同民族、不同民族国家之间存在显著的、实质性的差异。这种差异主要不是由生物学基因遗传决定的皮肤、眼睛颜色、身高等生物学性状产生的,而是由 “文化性状” 产生。考虑文化性状的人类群体,我们称为 “人类文化群体”(Human Cultural Group,HCG)。人类文化群体是由语言、习俗、文化传统、制度、法律、宗教信仰等主要决定因素界定的个体集合。这些决定因素是 “原因”,那么表现为 “结果”,就是群体性状。这样的群体性状表现为人口数量、经济发展水平、种群层面的领导力和影响力、军事势力等,一个简单的指标,例如是 “作为留学目的地” 所考虑、观察的那些指标。人类进化到现代,人类文化群体表现为民族国家,宗教团体等,主要是民族国家。考虑到定义人类文化群体的主要决定因素,韩国和朝鲜是两个不同的人类文化群体。
(二)复制者 - 互动者 - 群体性状概念框架
如果我们把人类文化群体视为一个选择单位,就像个体一样的一个实质性的事实存在,那么我们用复制者 - 互动者概念框架来刻画它。
互动者是一个部落、民族、民族国家或宗教群体 (例如凝聚力较强的犹太教群体)。
复制者是界定和区分一个人类文化群体与其他文化群体的作为 “原因” 的那些因素,如语言、习俗、文化传统、制度、法律、宗教信仰等。
群体性状是界定和区分一个人类文化群体与其他群体的作为 “结果” 的那些指标,例如人口数量、经济发展水平、种群层面的领导力和影响力、军事势力等。
人类文化群体的复制者的复制。复制者在人类文化群体的一代一代人之间复制,这种复制不必是 100% 高保真,但又不至于在几代之内变得不可识别其连续性,一个民族国家的语言、文化传统、基本法律和宗教信仰具有可遗传性。
文化群体之间的阻隔与交流。群体选择强调的是自然选择机制操作在许多群体之间,显然,每个群体之间的关系,就像只考虑群体内的个体竞争争夺食物和配偶一样,群体间存在竞争关系,它们之间的实力、规则制定领导的竞争。文化群体之间的边界清晰和群体间隔离,但群体间存在有限移民和群体间偶尔的征服 - 被征服;因为群体适应度持续降低,文化群体存在有限个数的灭绝。
(三)利己与利他的实质:非合作和合作
正如 Handley 和 Mathew (2020) 以肯尼亚北部 6 个部落 49 项社会规范的经验研究中发现,文化群体选择不必偏爱利他主义形式的合作行为以预测合作的社会规模和文化差异之间的对应关系。如果 CGS 影响到治理合作的规范,那么遵守这些规范符合个人的利益,因为偏离规范将导致社会不赞成和制裁。经济学家也有类似的理解,Jeroen 等 (2009) 指出,群体选择需要相互依赖的个体性实体,如非加性遗传或文化互动,或 “虑他行为”(other - regarding behavior)—— 不仅是利他行为,还有嫉妒、地位追求或任何其他社会互动。
我们认为,利己、利他概念具有误导性,它犯了 “直观 - 近因谬误”,为了修正这个谬误,我们提出:“合作行为” 替换 “利他主义行为”:合作性行为是指遵循群体内规则的,有保证 “复制” 过程顺利完成的 “顺从性” 行为。与合作行为相对的是 “单干”,合作行为是专业化 - 分工以创造合作剩余的行为,合作行为本身此后进一步进化到专业化 - 分工 - 非人格化交换的行为。合作行为的频率把个体行为 “结构化” 为群体性状,构成群体间合作剩余差异的直接原因。“顺从性” 的实质,是人类在社会影响或社会压力下对某种程序的易感性 (西蒙,2016:65;西蒙,2009:44);按照 “社会认可的方式行事” 并 “克制社会不认可行为” 的行为倾向 (西蒙,2016:76)。顺从性是从个体层级到群体层级转换的必要条件。因为文化和全部人工制品的代际复制对于文明是持续存在至关重要的,文化群体选择压力青睐顺从性,从而使顺从性成为人的基本行为倾向。顺从性并不假设 100% 的复制,事实也是如此,因此为行为准则为更一般 “文化”“变异” 留下了空间。给定人类个体以下三个特性:有限理性、高度依赖、文化继承 (这意味着一个远超个体经验容量的群体知识存贮量),那么群体选择压力青睐 “顺从性” 倾向和行为。人具有高度 “表型可塑” 性,顺从的天性使表型可塑成为可能。不过,过度的 “顺从性” 也可能使经济社会锁定在某种低效率状态。
“非合作性行为” 替换 “利己主义行为”(单干者):非合作性行为是指只考虑自己的 (个体的全部生物遗传和文化继承、理性能力叠加作用于个体,个体感知成本收益,计算成本收益,采取行动) 的行为倾向;遵循规则 (rule - following)、顺从 (docility) 等结构化力量不进入非合作性行为中。因高频率的非合作性行为导致合作剩余均值低于相对低频率的合作性行为群的合作剩余均值,所以群体选择导致非合作性行为降低的趋势,仅当维持群体隔离的力量强大时,非合作性行为频率维持一定稳态值。
| 理性条件 | 传统概念 | 修正概念 1 | 修正概念 2 | 演化稳定性 |
|---|---|---|---|---|
| 仅指涉自我 | 利己主义行为 | 非合作性行为 | 单干 (“百事通”) 策略 | |
| 指涉自我和他人 / 社会规范 | 利他主义行为 | 合作性行为 | 专业化 - 分工策略 | 演化稳定策略,ESS |
七、结论与展望
汉密尔顿法则试图按着基因选择的路线扩展到利他主义、专业化和分工 / 合作现象,它处理的是单一层级内的自然选择,基因选择或个体选择,因而回避了多层级选择 (群体选择),也就不能解释个体间结构化形成 “性状群体” 的多层级选择现象。在非人类动物世界是否存在群体选择有很大争议;但是在人类社会中,群体选择至少是自现代智人以后 (脑容量不再有增长的迹象) 驱动进化的主要力量,而在文化演化中,则是驱动进化的主导甚至是唯一力量。
从群内个体层级选择到群体层级的群体选择的跨越是一个 “涌现过程”,具有适应性优势的群体性状具有 “涌现性”,其中 “单干” 条件下不存在而结构化个体 (群体) 存在的巨大 “合作剩余” 正是群体性状的表征。群体性状的 “涌现性质” 摆脱了传统上自利与利他两难权衡问题,这种群体选择层级的涌现性状不需要来源于个体选择中的自我牺牲行为,而只需要遵循导向合作的一般抽象规则,而且这里的 “合作” 也不需要是个体有意识地与别人 “合作”,它的实质是专业化和交换。这样就大大松弛了传统上群体选择理论对 “利他主义” 的强依赖。基于如此理解的群体选择,我们将 “自利” 和 “利他” 置换为更符合涌现解释的 “单干策略” 和 “合作策略”(专业化分工和非人格化交换),而这样的重新界定并不影响普莱斯方程作为群体选择的基准解释模型。这里的合作策略是指通过规则将非血缘个体结构化,因此群体选择的选择单位就被简化为不同群体遵循的规则集。如此一来,被重新解释的普莱斯方程将不需要对维持一定频率的 “利他主义者” 来解释演化稳定均衡,而是诉诸决定群体性状的规则集的相对优劣势。
我们将传统演化理论中作为观察结果的 “繁殖成功度” 置换为不同群体遵循的 “规则集” 所覆盖人口数量。这不仅因为考虑到文化演化中的群体选择存在战争、征服、移民等 “非繁殖” 人口增量,更是考虑文化演化中群体选择作为观察结果的变量本身就不能从基因进化中的 “繁殖成功度” 移植过来,而有必要确立适用于文化演化领域自然选择的观察结果 ——“规则集覆盖人口数量”。
群体选择理论在经济学中的应用还是初步的。目前经济学中已经运用群体选择理论的议题有组织惯例的选择、组织结构的选择、群体选择在公共池塘资源困境中的作用,用群体选择理解社会经济权力概念,通过群体选择研究制度变迁和制度演化繁荣等。群体选择理论的关键预期 —— 群体选择导致大范围匿名个体间合作 (规则、执行机制、利他行为的先天心理倾向) 已经得到检验,在未来的研究中,除了以早期人类社会、当代世界中未开化部落为背景的经验以外,以现代社会为背景的企业间竞争、民族国家间竞争、跨国自由贸易协定联盟形成和分化、非经济目的的国际组织间竞争都可能通过群体选择理论得出有启发性的新发现。
社会合作秩序何以可能:社会科学的基本问题
韦倩 姜树广
内容提要:无亲缘关系的社会成员之间的大规模合作构成了人类社会的主要特征。一个显而易见的问题是:社会合作秩序得以维持的原因是什么呢?或者说,与其它物种相比,人类具有什么样的独一无二的特征才导致大规模群体中陌生人之间的合作呢?这个问题几乎是所有社会科学都关心的问题,但是,这个问题目前为止仍然存在一些未解之谜,仅仅依靠单一学科很难回答这个问题。本文对这个问题进行了详细的探索和深层次的追问,试图以跨学科的视角对这一社会科学的基本问题给出合理的解释。
关键词:社会秩序;合作;一阶搭便车;二阶搭便车;神经元经济学
与其他物种不同,即使在规模很大的群体中,人类也可以与非亲属成员进行合作(Boyd et al., 2003; Gintis, 2000)。这种无亲缘关系的社会成员之间的广泛合作构成了人类社会的主要特征。那么,社会合作秩序得以维持的原因是什么呢?或者说,与其它物种相比,人类具有什么样的独一无二的特征才导致大规模群体中陌生人之间的合作呢?这些问题几乎是所有社会科学都关心的问题,经济学、社会学、生物学、人类学等学科都对其给予了大量关注,并提出许多理论解释。但是,这些解释目前为止仍然存在一些问题和未解之谜,仅仅依靠单一学科很难回答这个问题。《科学》杂志在创刊 125 周年之际曾组织全世界最优秀的科学家提出了 125 个 “驱动基础科学研究以及决定未来科学研究方向” 的科学难题(其中 25 个 “大问题”,100 个 “小问题”),25 个 “大问题” 中有 23 个是纯粹的自然科学问题,只有两个问题同时还是社会科学问题,一个是 “马尔萨斯的人口论观点会继续错下去吗”,一个是 “人类合作行为如何演进”(黄少安和张苏,2013)。Pennisi(2005)在同期的《科学》杂志上指出,人类合作演进的问题是一个需要演化生物学家、动物学家、神经科学家、经济学家共同努力来研究的课题,需要最终探索出 “到底是什么因素在促成我们的合作精神”。英国皇家学会主席 May 在 2005 年的主席致辞中也曾说道:“人类社会和其他动物种群中的合作行为如何演化和维持是演化生物学和更广泛的社会科学中最重要的未解之谜”(Colman, 2006)。在本文中,我们将对这个问题进行详细的探索和深层次的追问,试图以跨学科的视角对这些问题和谜团进行探索和分析并给出合理的解释。
一、“一阶搭便车” 之谜
按照进化论的观点,人类可以在大规模群体中与非亲属成员进行合作的现象令人困惑(Boyd et al., 2003; Gintis, 2000),因为合作具有利他性,实施合作行为需要合作者自己负担成本,但却可以使群体内的非亲属成员获益。由于背叛者无需付出任何成本就可以从合作者身上获益,因此,理性的社会成员会采取背叛行为,对合作者进行 “搭便车”。这种现象被称为 “一阶搭便车” 行为。如果没有其他机制的约束,“一阶搭便车” 现象会逐渐蔓延并最终瓦解整个社会。那么,人类是如何避免 “一阶搭便车” 现象从而保持社会的合作秩序呢?除了同样适用于动物界的亲缘选择理论(Hamilton, 1964)外,学者们还提出了重复互动、声誉、群体选择与空间选择等理论。
一些学者认识到,很多社会交往是重复发生的,这或许能够解释存在于亲缘关系之外的合作行为。其代表性理论为互惠利他理论(reciprocal altruism theory)。该理论认为,如果未来重复交易的机会足够高,那么,在双边交易中,自然选择会偏爱合作行为(Trivers, 1971; Axelrod and Hamilton, 1981)。合作者在未来交易中收回合作的威胁会使得对手基于长远利益而抑制其背叛行为。与此相似,博弈论的研究成果也已表明,如果博弈双方再次相遇的概率比较大,折扣率也比较低,那么,即使参与者具有短期欺骗的动机,他们也会达成一个完全合作的均衡(Friedman, 1971; Fudenberg and Maskin, 1986)。
除此之外,社会秩序还经常表现为大量多边交易中的合作行为。基于此,一些学者提出声誉考虑的理论,比如间接互惠理论(indirect reciprocity)、有成本的信号理论(costly signaling theory)等。与互惠利他理论不同,间接互惠理论(Nowak and Sigmund, 1998; Wedekind and Milinski, 2000; Leimar and Hammerstein, 2001)的理念是:个体之所以选择合作行为是期望以此获得良好声誉,从而今后能得到第三方的奖励,而不是仅来自受惠个体的回报。Nowak 和 Sigmund(1998)考察了间接互惠的演化稳定条件,他们发现,如果合作行为的成本收益率小于行为主体获知受惠者类型的概率,那么,建立在间接互惠基础上的合作均衡将会非常稳固。与间接互惠理论不同,有成本的信号理论认为,通过合作行为,人们可以传递其作为潜在伙伴的信号,这使他们具有结盟优势,从而导致合作在无亲缘关系的社会群体中成功演化(Zahavi, 1995; Wright, 1999; Gintis et al., 2001)。比如 Gintis et al.(2001)通过构建一个以多人公共物品博弈作为分析框架(即多个发信号者 - 多个观察者)的理论模型发现,由于合作行为可以显示出发信号者的潜在品质,从而引出观察者的一个响应,这会使双方都受益,从而该行为是演化稳定的,在满足一定的条件下,尽管这种发信号行为起初非常稀少,但却可以在人口中增殖。
由于生物的进化不但可以在个体层次上实现,也可以在种群层次上实现,因此,一些学者(Henrich, 2004; Boyd and Richerson, 1985; Cavalli - Sforza and Feldman, 1981)提出了文化群体选择理论(cultural group selection theory)来解释社会合作秩序的维持。他们认为,当个体的合作行为有利于整个种群时,合作程度更高的群体会因适应性强而生存下来,从而合作的特征就可能随种群的保存而得以繁衍和保留。
另外一些学者注意到了社会交往的网络结构特征对群体合作秩序的影响,比如空间选择(spatial selection)或网络互惠(network reciprocity)理论认为,社会交往存在空间结构或网络差异,一些人的交往显然要比其他人更为紧密,这种结构差异会使得合作者通过空间聚集来避免被背叛者剥夺,从而空间选择效应可以促进合作的演化(Nowak and May, 1992; Ohtsuki et al., 2006)。比如 Ohtsuki et al.(2006)发现,如果合作或利他性行为的成本收益率超过相邻个体的平均数目,空间选择就会偏好合作行为。
令人遗憾地是,重复互动、声誉、群体选择与空间选择等任何一种机制都不允许利他性合作在规模很大的群体中演化。比如 Joshi(1987)、Boyd and Richerson(1988)等已经论证,条件合作的针锋相对之类的策略只能在 4 - 8 人的小规模群体中才能够成功演化,即使再次相遇的概率足够大,仅仅依靠针锋相对之类的策略或其他互惠行为也很难维持大规模群体的合作秩序。可见,重复交往只能支持一定规模内的二元关系之间的合作。而声誉、群体选择与空间选择机制只有在群体规模比较小且迁移很少发生的条件下才能导致合作行为的稳定演化(Leimar and Hammerstein, 2001; Rogers, 1990),比如 Leimar and Hammerstein(2001)发现,当群体发生混合时,合作率会急剧下降并逐渐趋近于零。
可见,上述理论均不能完美解释社会合作秩序何以可能的问题。一些学者相信,依靠惩罚威胁也许可以解决这个问题。由于背叛可能会遭受严厉的惩罚,使得背叛行为的成本大于合作的成本,这使得合作而不是背叛成为理性选择策略。大量实验(Fehr and Gächter, 2002; Fehr and Fischbacher, 2003; Camerer and Fehr, 2006)和人类学(Boehm, 1993; Henrich, 2004)的证据表明,许多人愿意自己承担成本去惩罚背叛者,甚至在一次性交往情况下也是如此。因此,惩罚可能是人类维持社会合作秩序的主要机制,惩罚行为的存在可以有效确立社会合作秩序(Gächter, et al., 2008; Henrich et al., 2006; Henrich and Boyd, 2001; Boyd et al., 2003)。这似乎表明,援引惩罚机制可以解决上述困惑,然而,非常遗憾,它却带来了一个新的演化之谜,下面我们将论述并尝试解释这个二阶搭便车之谜。
二、 “二阶搭便车” 之谜
虽然对背叛进行惩罚可以解决 “一阶搭便车” 问题,可是,惩罚需要付出成本,使得惩罚者的适应性降低。于是,纯粹合作者相对于惩罚性合作者来说就是一个 “二阶搭便车者(second - order free riders)”(Panchanathan and Boyd, 2004; Fowler, 2005),这样,就产生了一个新的演化之谜:由于惩罚者相对于二阶搭便车者来说需要负担额外成本,这必然使其在进化中丧失优势而消亡,那么,减少个人适应性的惩罚行为如何能通过自然选择得以进化呢?
Gintis(2000)认为,人类群体会以一定正的概率面临灭绝的威胁,比如战争、饥荒、自然灾害等,当人类面对灭绝的威胁时,由于未来太过渺茫,互惠利他和间接互惠都不能维持可以帮助群体幸存所必需的合作,并且大部分人类群体不仅仅局限于亲缘关系,使得亲缘选择在这里也不适合,但是,具有一定利他性惩罚者(也称强互惠者)的群体则可以在这种环境下幸存。利他性惩罚者自己承担成本而使整个群体受益,这虽然使得群内选择的演化压力不利于利他性惩罚行为,但却使得群间选择的压力更偏爱利他性惩罚行为,含有一定利他惩罚者的群体在这种情况下可以更好的生存。可见,利他性惩罚是一种演化上的适应反应,它可以给拥有它的族群带来可观的利益,并且该利益会超出个体为利他性惩罚行为所付出的成本。
Bowles and Gintis(2004)也从群体选择的角度去探讨二阶搭便车问题,但他们探究了一个不同的机制。在他们的模型中,发信号或群体灭绝都不起作用,惩罚采取驱逐(ostracism)和避开(shunning)的方式。在这种情况下,虽然利他性惩罚者承担了惩罚成本从而比其他人适存度降低,但是当群体成员都从共同遵守社会规范中获益时,他们也许会服从规范并惩罚违反规范者。他们使用系统动力学方法模拟了距今 10 - 20 万年以前更新世(pleistocene)晚期以狩猎 - 采集为生的人类社会环境,仿真结果表明,由突变产生较小数量的利他性惩罚者可以侵入自私者人群,使族群内的合作行为与适应性维持在一个较高水平。他们还运用 Price 方程实证检验了人口层次的力量会阻止合作者把利他性惩罚者驱逐出去。合作者由于是 “消极利他主义者”,虽然在群体内的适存度相对利他性惩罚者占优,但这个优势被大量合作者导致的群体较低平均适存度所抵消。如果群体效应足够大,那么合作者就不会把总人口中的利他性惩罚者驱逐出去,即便他们可以把利他性惩罚者从与他们共存的群体里赶走。
Boyd et al.(2003)给出了另外一种解释:尽管利他性合作和利他性惩罚的逻辑基础相似,但它们的演化逻辑却不相同,利他性合作者相对于背叛者的支付劣势独立于人口中背叛者的频率,但利他性惩罚行为的成本劣势却会随背叛者人数的下降而下降,当惩罚行为普遍时,群内反对惩罚的选择性压力相当微弱。正是这种非对称性的机制可以支持利他性惩罚在大规模群体中演化。他们的模拟结果显示,当没有惩罚时,群体选择只有在小规模情况时才能维持高频率的合作,而加上惩罚条件后,便可以在更大规模的群体中维持高频率的合作,且提高灭绝率可以提高长期平均的合作水平;另外,长期平均合作水平对惩罚成本非常敏感,当惩罚成本较大时,群间选择便不能补偿群内惩罚者的下降,最终使得惩罚者和捐献者都会减少;当背叛者减少时,只有在惩罚者的支付成本可以随之降低的情况下,惩罚才可以导致合作的提高,而如果惩罚者承担的成本固定,即独立于群体中背叛者的人数,则惩罚不会对合作的演化有所帮助。他们的模型揭示了不能维持利他性合作的群间选择却可以产生利他性惩罚演化的可能。
与上述研究不同,Henrich and Boyd(2001)的社会顺从(conformism)理论认为,合作和惩罚的演化是人类在同化过程中采用流行行为的一个副效应,文化演化模型可以解释一次交往情境中的惩罚行为。他们认为,人类的社会学习并不是随机复制别人的行为,而是复制成功者和大多数人的行为。这种学习机制虽然不能总是产生最优行为,但却可以快速地获得高适应行为,正是这种收益偏好传播和从众传播机制使得社会可以建立合作和惩罚的规范。他们的文化演化模型显示,在有限次惩罚阶段的条件下,仅有少数尊奉习俗者的传播就可以在演化均衡中稳定群体的合作和惩罚行为。一旦一个群体的合作稳定后,便可以通过群体选择机制使得群体受益的文化特征在一个多群体的人口中扩散。文化群体选择进程主要以两种方式来扩散一些亲社会行为,一是合作的群体具有更高的生产能力,从而可以支持更快的人口增长;二是模仿成功人士的行为也会使得合作行为被模仿。最后,一旦合作均衡变得普遍时,自然选择也会偏爱那些喜欢合作和惩罚背叛者的基因,因为这些基因降低了人们遭受惩罚的成本。
另有一些学者注意到,人类社会中存在各种社会排斥(social exclusion)机制,比如驱逐、躲避和忽视等,这会使群体中并非所有成员都能享受合作或惩罚行为的益处,从而有利于群体合作秩序的演化。这种观点被称为社会排斥理论。Panchanathan and Boyd(2004)构建了一个包含间接互惠机制的互助游戏环境,其中,回避者(Shunner)只帮助具有好名声的弱者,而不会为坏名声者提供帮助。他们发现,这种基于声誉的社会排斥机制具有演化上的稳定性,可以提供对有成本合作行为的激励。同样,利他性惩罚者也可以通过排斥来约束搭便车者参与利益分享。Sasaki and Uchida(2013)的博弈理论模型表明,排斥可以减少合作受益者的数目,从而增加其利益分享,由于驱逐者的收益超过被驱逐的背叛者,只要驱逐的成本比较低,即使在背叛者比较多的群体驱逐行为也会占优而得到扩散。这表明,社会排斥可以有效克服二阶搭便车问题。
还有一些学者注意到,社会交往的空间结构特征也会对惩罚的扩散有影响,当它可以促进惩罚行为在群体中的扩散时,就会淘汰二阶搭便车者。Helbing et al.(2010)认为,与利他性惩罚者不同,人群中还存在另一类惩罚者,虽然他们自身也是背叛者,但却会对背叛者进行惩罚,这类人被称为伪善者或背德者。在可与邻近个体进行空间交往的情形下,利他性惩罚者可以通过空间分割完全驱除搭便车者,虽然他们起初数量会严重下降,但最终会扩散到整个人群;利他性惩罚者还可以从与伪善者的罪恶同盟(unholy cooperation)中获益。不同于 Helbing et al.(2010)的伪善者,Szolnoki and Perc(2013)在空间交往情景中引入了有条件的惩罚者以区别于一般模型中的无条件惩罚者,有条件惩罚者的惩罚程度与其他惩罚者的数量成比例。他们发现,当惩罚成本较高时(更符合现实的情况),有条件惩罚比无条件惩罚更能有效维持合作效率。
还有其他一些非常有价值的研究也对二阶搭便车难题提供了部分解决方案,如多重惩罚(Boyd and Richerson, 1992)、变异(Sethi and Somanathan, 1996)、公共品的回报额度(叶航,2012)等,不再一一而述。
虽然上述研究从不同角度对 “二阶搭便车问题” 进行了解释,提出了非常有价值的洞见,但却仍然存在一个明显的未解之谜。它们均假设惩罚者能够低成本实施严厉惩罚,即惩罚者付出的成本要小于惩罚对背叛者造成的伤害,只有这样的条件下,惩罚和合作行为才能够成功演化。这个假设在一定程度上是合理的,因为人类具有高超的制造工具和狩猎的能力,比如投掷抛射物等,从而是对背叛者可以低成本施加严厉处罚的唯一物种(Bingham, 1999);或者小数目的惩罚者也可以通过协商、秘密行动和致命武器等手段来降低适应性成本从而击败最强大的单个敌人(Gintis, 2000)。但是,这些能力显然不只惩罚者独有,这使得惩罚者有时不是背叛者的对手,因此,低成本惩罚的假设并不总令人信服。另外,人们对惩罚的反应并非总是容忍,报复非常常见,这也会加大惩罚实施成本,从而降低了利他性惩罚理论的可信性与适存度。比如:Janssen 和 Bushman(2008)经过模拟发现,如果报复行为被允许,合作和利他性惩罚不会在大规模群体中演化。很明显,如果惩罚者不能低成本实施严厉惩罚,那么,依靠惩罚来维持社会合作秩序的信念将很难再被坚持。可见,惩罚强度和惩罚成本这两个变量对合作的演化之谜起到至关重要的作用。下面我们通过仿真分析来表明这点。
该软件的详细介绍参见浙江大学叶航教授的个人主页:
http://mypage.zju.edu.cn/yehang
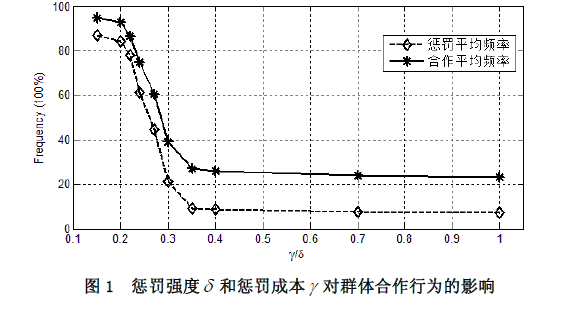
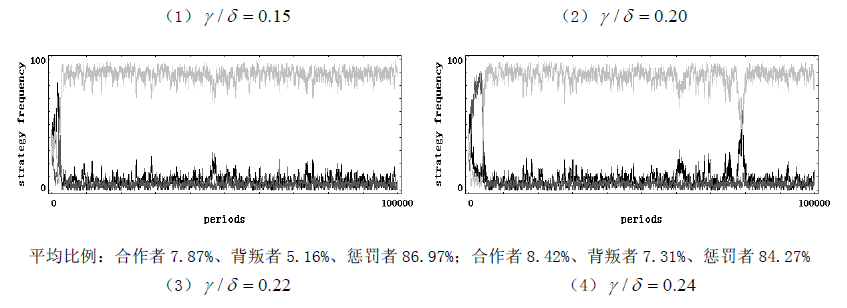
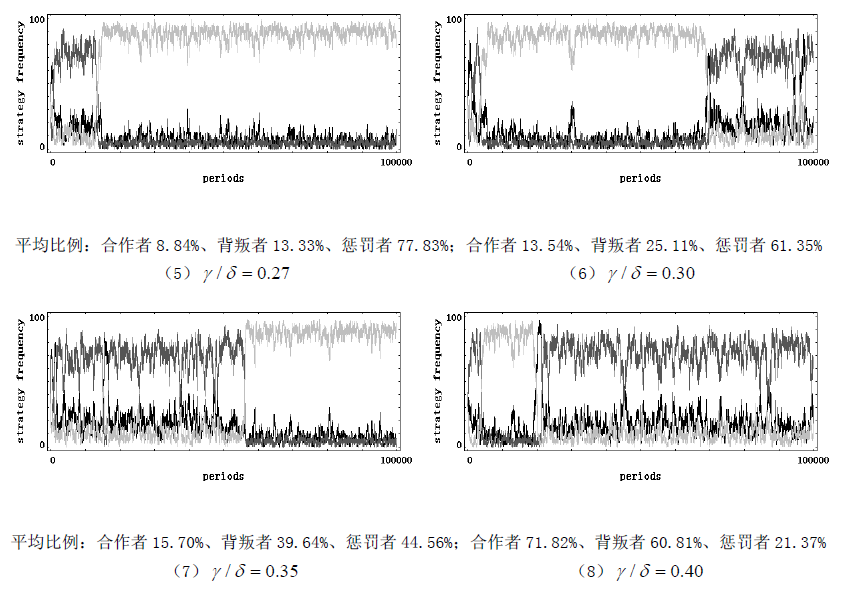
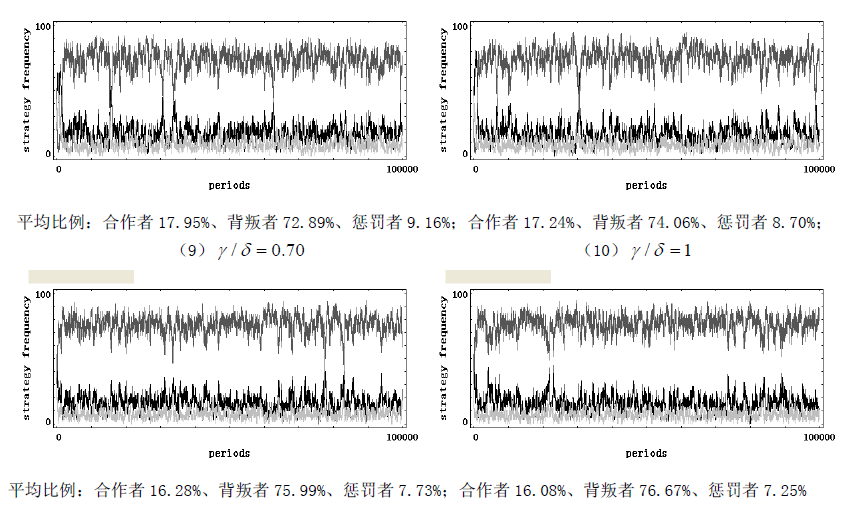
考虑一个规模恒为 m m m 的人群,每次随机抽取 I I II II 个成员参加一个公共物品博弈,假定每个成员都有三种行为选择 —— 只捐献不惩罚(合作者)、不捐献(背叛者)、既捐献又惩罚(惩罚者)。假设该群体中合作者的数量为 x x x,背叛者的数量为 y y y,惩罚者的数量为 z z z,那么一定有 x + y + z = n x + y + z = n x+y+z=n。捐献需要个人支付成本 c c c,但会产生一个可以被全体成员均分的总收益 b = r c b = rc b=rc,其中 r > 1 r>1 r>1 为合作回报乘数;背叛虽然不需支付任何成本,但也不会给群体带来任何收益;惩罚会使群体内的每一个背叛者减少 δ \delta δ 的收益,但也会给惩罚者带来 y y y 的成本。不考虑同情以及监视成本等其他因素,可知:每轮博弈中捐献者的收益为 x + z n b − c \frac {x + z}{n} b - c nx+zb−c,背叛者的收益为 x + z n b − δ z \frac {x + z}{n} b - \delta z nx+zb−δz,惩罚者的收益为 x + z n b − c − γ y \frac {x + z}{n} b - c - \gamma y nx+zb−c−γy。可以看出,惩罚相对于合作来说损失了 γ y \gamma y γy 的成本,理性的个体是不愿意付出惩罚的。但是,群间选择的力量却偏爱惩罚。假设个体的行为可以进行模仿,并且假定个体更倾向于模仿更高收益的行为。此外,还假定在每一轮博弈开始之前, m m m 中的所有成员都将以一个很小的概率 μ \mu μ 随机突变成其他类型。在完成这些步骤后,再从 m m m 中随机抽取 I I II II 个成员进行新一轮博弈。假定这个公共物品博弈的随机演化过程是一个频率依赖性的 Moran 过程。我们使用浙江大学跨学科社会科学研究中心(ICSS)开发的仿真软件 PGG - SPC++2.0 对这个模型进行了代数为 1 x 10 1x10 1x10 的仿真 ①(仿真具体结果参见附录),仿真支持了我们的观点。如图 1 所示,横轴表示惩罚成本与惩罚强度之比 γ / δ \gamma / \delta γ/δ,纵轴表示各行为在人群中的平均频率,随着 γ / δ \gamma / \delta γ/δ 的增大,惩罚者在人群中的平均比例逐渐下降,从而群体的合作程度也逐渐下降。主要参数的取值状况 ①为: γ / δ = ( 0.15 、 0.20 、 0.22 、 0.24 、 0.27 、 0.30 、 0.35 、 0.40 、 0.70 、 1.00 ) \gamma / \delta=(0.15、0.20、0.22、0.24、0.27、0.30、0.35、0.40、0.70、1.00) γ/δ=(0.15、0.20、0.22、0.24、0.27、0.30、0.35、0.40、0.70、1.00)、 m = 100 m = 100 m=100、 n = 5 n = 5 n=5、 x = 30 x = 30 x=30、 y = 40 y = 40 y=40、 z = 30 z = 30 z=30、 c = 1 c = 1 c=1、 r = 4 r = 4 r=4、 μ = 0.001 \mu = 0.001 μ=0.001。除此之外,我们还分别对初始状态( ( x , y , z ) (x, y, z) (x,y,z) 以及公共物品博弈的人数 n n n 进行了稳健性检验,仿真结果与图 1 完全相似,限于篇幅略去。
因此,一个值得进一步分析和探索的问题将是:族群或社会如何保证惩罚者能以低成本对背叛者进行严厉的惩罚?我们认为,族群中必然存在一些可以提高人类惩罚能力的机制,这有助于利他性惩罚行为在人群中稳定演化,并最终维持了群体的合作秩序。比如:一些学者认为惩罚行为并非总是无条件的或者是没有协调的个人自发行为,而是常常通过小道传闻或与其他惩罚者的沟通而得以协调进行(Wiessner,2005;Boehm,1993),因此,惩罚可以作为一种集体行为向背叛者传递群体的谴责信息。Boyd 等(2010)的模型就认为,协调的惩罚更符合人类祖先的现实条件,这会使得随着惩罚者人数增加,惩罚单个搭便车者的平均成本会降低,从而即使在惩罚者稀少的情况下,惩罚也会得到繁衍从而增加群体的收益;而自发的同辈惩罚(peer punishment)只在群体的规模足够小或个体有不合作的选择权利等限制条件下才有效。还有一些学者认为,群体具有趋向形成自我管理的机制,惩罚背叛者的权力一般被赋予特定的权威集中者,这可以提高惩罚的效力(Henrich 等,2010;Greif 等,1994)。比如传统社会的村民会请求他们的首领来调节争端(Gibson 和 Marks,1995)、中世纪的欧洲商人创立了工会来维持商业秩序(Greif,1993)等。在人类社会中,惩罚性的制度安排实际担当了维持社会秩序的主要责任。惩罚基金(punishment fund)可被看作维护公共利益的制度雏形,许多小规模的社会都使用这种方式(如通过雇佣一个执法者),如 Ostrom(1990)描述了许多自筹资金保证合约执行的例子。由于集权化的制度可以更有效的克服协调问题和二次搭便车问题,因此,从演化的角度来看比同辈惩罚更有效率(Güth 等,2007),可见,人类社会的层级结构和权威在合作秩序维护中扮演了重要的角色,甚至角色分化和等级统治也是大多数动物群体的组织原则。特别是在复杂的大规模社会,集权化惩罚(centralized sanctioning)和法定权威对维持社会的合作秩序更有意义,他们的权威来自所处社会阶层的位置(Baldassarri 和 Grossman,2011)。韦倩(2009)思考了人类社会中存在的提高惩罚能力的三种社会机制 —— 规范的内化、缔结同盟与第三方介入,分析了它们在人类社会中的作用机理和相关影响因素,并构建了一个动态演化模型探讨了这三种机制对于维持利他性惩罚行为的演化和社会合作秩序的作用。
可见,人类族群或社会中存在一些可以提高惩罚能力的社会机制,即使在单个惩罚者的惩罚能力不能够满足惩罚行为稳定演化的条件下,这些机制也会自动调整其惩罚能力,使得利他性惩罚行为能够在人群中保持稳定演化,从而维持了社会的合作秩序。因此,我们的研究可以大大增强利他性惩罚理论的解释力度和适用范围。
三、 合作行为的神经学基础
虽然利他性惩罚是维持社会合作秩序的主要机制,并且族群或社会中存在许多机制使利他性惩罚得以实施,从而避免了二阶搭便车问题,但是这相对于人类丰富多彩的合作行为来讲仍显不足,因为它并不一定可以保证惩罚行为对背叛者造成的伤害一定大于惩罚者付出的成本,即使高级形式的制度安排仍然面临执法者犯法的危险。
对不惩罚背叛者的人进行惩罚(二阶惩罚)也许可以完美解决 “二阶搭便车” 的问题(Boyd 和 Richerson,1992;Colman,2006),但是,这会带来更高阶的 “三阶搭便车”,依次类推,还可能存在四阶、五阶乃至无穷阶的搭便车问题。比如,集权化制度中的权威者也可能会监守自盗,以及即使在社会制度已经发育完善的现代社会中仍然存在无法被完全监督和制约的个体(如国家元首)等。现有文献对高阶搭便车现象的分析非常少见。运用逆推法可知,当搭便车的阶数 N N N 趋向于无穷时,由于不存在更高价的惩罚,此时再也不能依靠物质利益来约束搭便车者,这样,搭便车者会破坏这一层级内的合作;再推到前一阶,当成员预期到上一层级内的合作秩序不能维持时,他们也不会愿意自己承担成本去维持这一层级内的合作秩序,如此类推,最终整个社会体系都会崩溃,无论 N N N 的链条延长到何种程度都不管用。因此,当考虑到无穷阶搭便车问题时,再也不能从物质利益的角度来解释搭便车问题了,这时候社会秩序的维持和运转只能依靠社会成员内心的道德约束。那么,人类内心是否天生就具有愿意与别人合作、愿意自己付出成本惩罚别人、具有关心他人的亲社会情感呢?神经元经济学 ①的研究表明,人是被情感、情绪、道德等因素支配的综合社会人,人类心智结构的发展也体现出对合作秩序遵从的固有倾向。
首先,人类具有奖励合作行为的心智结构。人类之所以可以牺牲自身的物质利益参与合作,可能源自个体在相互合作中可以获得更高的快乐体验。比如 Rilling 等(2002)发现,被试在与人类对手相互合作时可以比他与电脑对手合作时获得更高程度的纹状体激活,由于在这两种情形下其货币收益是相同的,因此,这表明他从合作行为中获得了额外收益。但这个建立在重复博弈之上的实验可能会混同其他影响因素,从而影响结果的可信性,因此,Rilling 等(2004)进行了关于一次序贯社会困境博弈的实验,实验结果再一次表明,与计算机对手情况相比,被试与人类对手的合作可以产生更高的纹状体激活。Singer 等(2004)完成了一个更深层次的实验,他们发现,仅仅看到以前合作的个体的脸就可以激活激励相关脑区,这揭示了合作行为的特殊效用品质。这个结果意味着人们可以从与合作的人们的相互交往中获得更大的效用,这不仅是因为他们在这些互动中可以获得更多的金钱,而是因为这些互动本身就具有奖励作用。
其次,人类具有奖励惩罚行为的心智结构。神经与认知科学的证据表明,观察到背叛行为没有得到惩罚是一件痛苦的事情,同时人们从惩罚背叛行为本身中也可以获得效用的满足。De Quervain 等(2004)利用正电子发射成像技术对二人序贯社会困境博弈进行的研究对此提供了充分的神经学证据。由于大脑的背侧纹状体包括尾核与壳核的神经回路是人类及灵长类动物的一个与激励相关的关键部位,因此,当对背叛进行惩罚时如果能观察到这一脑区的激活,那么,就可以证实人们能从惩罚本身中获得收益。在被试 A 将自己的货币全部交给 B 后而 B 选择什么也不给 A 的情况下,实验者为 A 设计了 4 个不同场景:(1) I C IC IC,即惩罚对于 A 和 B 都是有代价的,此时 A 施加给 B 的每一个惩罚点将会给 A 带来一单位代币的成本,但却可以让 B 损失两单位代币;(2) I F IF IF,即惩罚对于 B 有代价但是 A 没有代价,此时 A 花费一个惩罚点来惩罚 B 时自己没有任何成本但是会让 B 损失两单位货币;(3) I S IS IS,即 A 对 B 的惩罚是象征性的,此时,惩罚既不能减少 A 的货币也不能减少 B 的货币;(4) N C NC NC,即 B 的行为是随机的且 A 事先知道,但是惩罚对于 A 和 B 来说仍然是有代价的。实验结果显示,A 在 I C IC IC、 I F IF IF 和 I S IS IS 三种情况下都表现出强烈的惩罚愿望:在 I F IF IF 条件下,全体被试都对 B 实施了惩罚;在 I C IC IC 条件下,14 个被试中有 12 个对 B 实施了惩罚;在 I S IS IS 条件下,14 个被试中有 6 个对 B 实施了惩罚。与此形成鲜明对比的是,在 N C NC NC 条件下,A 几乎没有惩罚愿望,14 个被试只有 3 个惩罚了 B,而且惩罚强度相当低。这个实验揭示了人们宁愿舍弃私利也要惩罚背叛者的动力所在,即惩罚者可以从惩罚行为本身中获得欲望满足。在另一项研究中,Singer 等(2006)在被试或其搭档被电击(轻微疼痛)时使用 fMRI 扫描其大脑发现,当合作的搭档被电击时,被试的前扣带区和前脑岛都观察到移情反应(empathic responses),而伏核(NAcc)和 OFC 区域则在不合作者被电击时表现活跃,被试报复动机的强烈程度也与 NAcc 的活跃程度相关,这表明对不合作者惩罚可以得到奖赏。
再次,人类具有关爱他人的心智结构。这种心智结构可能起源于哺育后代的挑战(Hrdy,2011)或来自种群内的冲突(Bowles 和 Gintis,2011)。最近有关大脑神经系统的一些研究显示,人们生来就具备关心他人的神经系统 —— 迷走神经(vagus nerve)。迷走神经位于胸部,从脊髓顶端开始,然后在全身漫游,分布于面部肌肉组织、与发音相关的肌肉、心脏、肺、肾脏、肝脏以及各种消化器官等,主管人的发声、心脏和呼吸节律、消化和其他功能。克特纳(2009)认为,当人产生同情时,迷走神经会处于最兴奋的状态,这会让人作出牺牲。Porges(2011)认为,迷走神经通过以下三种方式产生怜悯之情:首先,迷走神经可以调动参与关爱行为的各个肌肉群,包括面部肌肉组织和发音器官等。比如当一个人听到受苦受难的描绘时,往往会发出有节奏地叹息。这声叹息是关切与理解的情感表达,它会使叹息者平静下来,并可以缓解描绘者的痛苦,增加其信任。其次,迷走神经是控制人类心率的首要制动装置。如果迷走神经没有被激活,人的心脏平均要跳动 115 次,而不是一般情况下的 72 次。当人们愤怒或害怕时,心跳就会加快,把血液输送到各肌肉群,让身体做好战斗或逃跑准备。迷走神经的作用正好相反,它可以降低心率,让人心平气和,增加了与他人近距离温柔接触的可能性。再次,当迷走神经启动时,会引发后叶催产素的释放,而催产素是引致亲社会行为的一种激素,它会在大脑和全身各处发出了温暖、信任、热爱的信号,刺激出现了亲切友好的说话腔调以及更为平静的心血管生理现象,这些信号最终还会传达到别人身上。可见,迷走神经经常是各种亲社会行为的来源。值得一提的是,迷走神经是哺乳动物所特有的神经,或许正是它支撑了哺乳动物特殊的照管后代行为。
最后,人类具有同情共感的心智结构。迷走神经只可以激发人的 “同情”,但是个体如何感知别人的痛苦、快乐呢?这就要依靠镜像神经元。神经与认知科学的研究发现,包括人在内的灵长类生物的大脑中央运动前皮层有一个 F 5 F5 F5 区域,该区域的功能与其他绝大多数神经元不同,大多数神经元的活跃程度是与特定的动作联系在一起的,而这些神经元的活动并不与任何简单的、构成一个完整动作的 “元动作” 相关联,而是与一个完整的动作相关联(Rustichini,2005)。镜像神经元的激活具有以下特征:首先,当它们观察到一个特定动作时,就会作出反应,且与这个动作主体相距的距离无关;其次,镜像神经元是对动作,而不是对动作的对象作出反应;再次,镜像神经元是对动作在内心的呈现产生反应,而不是针对动作的物理现实,比如 Umiltá 等(2001)通过实验证实,激活神经元需要完整的动作,而且这与动作是否有一部分被隐藏而不能被观察到无关;最后,内在心理状态也存在镜像,Wickers 等(2003)通过实验证实,被试观察到的情绪也会激活其相关的 “镜像神经元”,因此,这表明人们亲身感受某种情绪时所激活的脑区,与他观察到其他人感受同一种情绪时所激活的脑区确实存在着重叠部分(Rustichini,2005)。可见,同情具有大脑活动的基础,“同情” 只不过是做某件事和观察到同一件事时基于 “镜像神经元” 实现的 “神经网络共享”。在人类具有了同情共感的能力后,才可以将制度内化于心。
可见,正是人类天生具有愿意与别人合作、愿意自己付出成本惩罚别人、具有关心他人的亲社会情感的禀赋,社会秩序才得以维持。最近的一项研究也表明,合作在很大程度上由直觉神经系统实现而不是由慎思神经系统来完成的。由于直觉神经系统的决策时间一般较短,而慎思神经系统的决策时间相对较长,因此,Rand 等(2012)通过公共物品实验研究被试的决策时间与最终捐献率之间的相关性,他们发现决策时间短的被试捐献得更多,从而更倾向于合作;而且当要求被试加快决策速度时可以提高其捐献率,当提示他们放慢决策速度时会降低其捐献率。可见,合作在很大程度上不是算计的结果,而是人类天生的心智结构自动完成的。对照我们日常生活中的行为决策,我们每个人都应该对此有所感知。当然,这里还存在一个问题:人类是如何演化出这样的心智结构呢?虽然这需要对人类原始社会的生活形态进行更深入的探察和更严谨的分析,但道理却可能非常简单:人类偏爱合作的心智结构是人类在长期演化过程中通过自然选择固化的产物,即自然选择会使得人类进化出偏好合作行为的神经机制,并把它编码到自动或情感的神经过程中。我们认为,一种很可能的途径是:原始人最初只是小规模的群居生活,日常交往对象都是亲戚和近邻,这时候亲缘选择和直接互惠的动机发挥主要作用,在这种情景下,人类出于哺育后代的压力等演化出同情共感、关爱他人、惩恶扬善的能力以及丰富的情感情绪,从而具有了在更大范围合作的神经基础。在此基础上,人类的交往与合作范围开始扩大,因为此时间接互惠以及利他性惩罚开始扮演维持合作秩序的主要角色,再加上人类社会已经演化出增强惩罚能力的一些机制,人类的生产水平也能够支持微弱劣势的个体不会立刻被演化淘汰,从而利他性惩罚者可以始终在人群中维持一定的比例,构成社会秩序的终极守护者,使人类成为自然界中唯一一个可以在大规模群体中与非亲属成员进行合作的物种。
四、 结论
与其他物种不同,即使在规模很大的群体中,人类也可以与非亲属成员进行合作。这种无亲缘关系的社会成员之间的广泛合作构成了人类社会的主要特征。那么,社会合作秩序得以维持的原因是什么呢?对人类合作难题的探究引发了学术界极大的兴趣。首先,学者们先注意到了 “一阶搭便车” 之谜,即背叛者对合作者的搭便车现象,并基于重复互动、声誉与群体选择等机制进行解释,但是,它们中的任何一种机制都不支持利他性合作行为在大规模群体中演化。于是,许多研究人员相信,依靠惩罚威胁可以维持人类大规模的合作。但它却带来了另外一个新的演化之谜,与惩罚性合作者相比,只合作而不惩罚者就成为一种 “二次搭便车者”。然后,本文对二阶搭便车问题的相关文献进行总结,对比分析了它们的不足和缺陷。还通过仿真讨论了惩罚强度和惩罚成本这两个变量对二次搭便车问题的影响,并讨论了人类社会上存在的一些可以改变它们的社会机制,发现即使单个惩罚者的惩罚能力不能够满足惩罚行为稳定演化的条件,一些社会机制也会自动调整其惩罚能力,使得利他性惩罚行为能够在人群中稳定演化,维持了群体合作秩序,这大大增强了利他性惩罚理论的解释力度和适用范围。或许对不惩罚背叛者的人进行惩罚可以完美解决 “二阶搭便车” 的问题,但是,这会带来更高阶的 “三阶搭便车” 问题,依次类推,还可能存在四阶、五阶乃至无穷阶的搭便车问题。“社会何以可能” 的问题最终需要向人的内心去追问。神经经济学的最新研究成果支持了我们的观点,神经经济学的研究表明,人类内心天生就具有愿意与别人合作、愿意自己付出成本惩罚别人、具有关心他人的亲社会情感的心智结构。
附录:
仿真图(参数的取值状况为: m = 100 m = 100 m=100、 n = 5 n = 5 n=5、 x = 30 x = 30 x=30、 y = 40 y = 40 y=40、 z = 30 z = 30 z=30、 c = 1 c = 1 c=1、 r = 4 r = 4 r=4、 α 1 = 1 \alpha_{1}=1 α1=1、 δ = 1 \delta = 1 δ=1、 ω = 1 \omega = 1 ω=1、 μ = 0.001 \mu = 0.001 μ=0.001),其中黑色线条表示合作者、褐色线条表示背叛者、浅色线条表示惩罚者。
参考文献
达契尔・克特纳,2009:《生而向善》,中国人民大学出版社。
黄少安和张苏,2013:《人类的合作及其演进:研究综述和评论》,《中国社会科学》第 7 期。
韦倩,2009:《增强惩罚能力的若干社会机制与群体合作秩序的维持》,《经济研究》第 10 期。
叶航,2012:《公共合作中的社会困境与社会正义》,《经济研究》第 8 期。
叶航、汪丁丁和贾拥民,2007:《科学与实证:一个基于 “神经元经济学” 的综述》,《经济研究》第 1 期。
Axelrod, R., and W. D. Hamilton, 1981, “The Evolution of Cooperation”, Science, 211: 1390 - 1396.
Baldassarri, D., and G. Grossman, 2011, “Centralized Sanctioning and Legitimate Authority Promote Cooperation in Humans”, Proceedings of the National Academy of Sciences of the United States of America, 108: 11023–11027.
Bingham, P. M., 1999, “Human Uniqueness: A General Theory”, Quarterly Review of Biology, 74 (2): 133 - 169.
Boehm, C. H., 1993, “Egalitarian Behavior and Reverse Dominance Hierarchy”, Current Anthropology, 34: 227 - 254.
Boyd, R., H. Gintis, S. Bowles, and P. J. Richerson, 2003, “The Evolution of Altruistic Punishment”, Proceedings of the National Academy of Sciences of the United States of America, 100 (6): 3531 - 3535.
Boyd R., and P. J. Richerson, 1985, Culture and the Evolutionary Process, Chicago: University of Chicago Press.
Boyd, R., and P. J. Richerson, 1988, “The Evolution of Reciprocity in Sizable Groups”, Journal of Theoretical Biology, 132: 337–356.
Boyd, R., and P. J. Richerson, 1992, “Punishment Allows the Evolution of Cooperation (or Anything Else) in Sizable Groups”, Ethology and Sociobiology, 13: 171 - 195.
Boyd, R., H. Gintis, and S. Bowles, 2010, “Coordinated Punishment of Defectors Sustains Cooperation and Can Proliferate When Rare”, Science, 328: 617–620.
Bowles, S., and H. Gintis, 2004, “The Evolution of Strong Reciprocity: Cooperation in Heterogeneous Populations”, Theoretical Population Biology, 65: 17–28.
Bowles, S., Gintis, H., 2011. A Cooperative Species: Human Reciprocity and its Evolution, Princeton, NJ: Princeton University Press.
Camerer, C. F., and E. Fehr, 2006, “When Does Economic Man Dominate Social Behavior?”, Science, 311: 47 - 52.
Cavalli - Sforza, L. L. and M. W. Feldman, 1981, Culture Transmission and Evolution, Princeton, NJ: Princeton University Press.
Colman, A. M., 2006, “The Puzzle of Cooperation”, Nature, 440: 744–745.
De Quervain Dominique J. - F., U. Fischbacher, V. Treyer, M. Schellhammer, U. Schnyder, A. Buck and E. Fehr, 2004, “The Neural Basis of Altruistic Punishment”, Science, 305: 1254 - 1258.
Fehr, E., and S. Gächter, 2002, “Altruistic Punishment in Humans”, Nature, 415: 137 - 140.
Fehr, E., and U. Fischbacher, 2003, “The Nature of Human Altruism”, Nature, 425: 785–791.
Fowler, J. H., 2005, “Second - order Free - riding Problem Solved?”, Nature, 437 (7058): E8 - E8.
Friedman, J., 1971, “A Noncooperative Equilibrium for Supergames”, Review of Economic Studies, 38: 1 - 12.
Fudenberg, D., and E. Maskin, 1986, “The Folk Theorem in Repeated Games with Discounting or with Incomplete Information”, Econometrica, 54: 533 - 556.
Gächter, S., E. Renner, and M. Sefton, 2008, “The Long - Run Benefits of Punishment”, Science, 322 (5907): 1510–1512.
Gibson, C. C., S. A. Marks, 1995, “Transforming Rural Hunters into Conservationists: An Assessment of Community - Based Wildlife Management Programs in Africa”, World Development, 23: 941–957.
Gintis, H., 2000, “Strong Reciprocity and Human Sociality”, Journal of Theoretical Biology, 206: 169–179.
Gintis H., E. A. Smith and S. Bowles, 2001, “Costly Signaling and Cooperation”, Journal of Theoretical Biology, 213: 103 - 119.
Greif, A., 1993, “Contract Enforceability and Economic Institutions in Early Trade: The Maghribi Traders’ Coalition”, American Economic Review, 83: 525–548.
Greif, A., P. Milgrom, and B. R. Weingast, 1994, “Coordination, Commitment, and Enforcement: The Case of The Merchant Guild”, Journal of Political Economy, 102: 745–776.
Güth, W., M. V. Levati, M. Sutter, E. van der Heijden, 2007, “Leading by Example with and without Exclusion Power in Voluntary Contribution Experiments”, Journal of Public Economics, 91: 1023–1042.
Hamilton, W. D., 1964, “The Genetical Evolution of Social Behaviour, I and II”, Journal of Theoretical Biology, 7 (1): 1–52.
Helbing, D., A. Szolnoki, M. Perc, and G. Szabó, 2010, “Evolutionary Establishment of Moral and Double Moral Standards through Spatial Interactions”, PLoS Computational Biology, 6 (4): e1000758.
Henrich, J. P., 2004, Foundations of Human Sociality: Economic Experiments and Ethnographic Evidence From Fifteen Small - Scale Societies, Oxford: Oxford University Press.
Henrich, J., and R. Boyd, 2001, “Why People Punish Defectors: Weak Conformist Transmission can Stabilize Costly Enforcement of Norms in Cooperative Dilemmas”, Journal of Theoretical Biology, 208: 79 - 89.
Henrich, J., R. McElreath, A. Barr, J. Ensminger, C. Barrett, A. Bolyanatz, J. C. Cardenas, M. Gurven, E. Gwako, N. Henrich, C. Lesorogol, F. Marlowe, D. Tracer, and J. Ziker, 2006, “Costly Punishment across Human Societies”, Science, 312 (5781): 1767–1770.
Henrich, J., J. Ensminger, R. McElreath, A. Barr, C. Barrett, A. Bolyanatz, J. C. Cardenas, M. Gurven, E. Gwako, N. Henrich, C. Lesorogol, F. Marlowe, D. Tracer, and J. Ziker, 2010, “Markets, Religion, Community Size, and the Evolution of Fairness and Punishment”, Science, 327 (5972): 1480–1484.
Hrdy, S. B., 2011, Mothers and Others: The Evolutionary Origins of Mutual Understanding, Cambridge, MA: Harvard University Press.
Janssen, M. A., and C. Bushman, 2008, “Evolution of Cooperation and Altruistic Punishment When Retaliation Is Possible”, Journal of Theoretical Biology, 254: 541 - 545.
Joshi, N. V., 1987, “Evolution of Cooperation by Reciprocation within Structured Demes”, Journal of Genetics, 66 (1): 69–84.
Leimar, O., and P. Hammerstein, 2001, “Evolution of Cooperation through Indirect Reciprocity”, Royal Society, 268: 745 - 753.
Nowak, M. A., and K. Sigmund, 1998, “Evolution of Indirect Reciprocity by Image Scoring”, Nature, 393: 573 - 577.
Nowak, M. A., and R. M. May, 1992, “Evolutionary Games and Spatial Chaos”, Nature, 359 (6398): 826–829.
Ohtsuki H., C. Hauert, E. Lieberman, and M. A. Nowak, 2006, “A Simple Rule for the Evolution of Cooperation on Graphs and Social Networks”, Nature, 441: 502 - 505.
Ostrom, Elinor, 1990, Governing the Commons, New York: Cambridge University Press.
Panchanathan, K., and R. Boyd, 2004, “Indirect Reciprocity Can Stabilize Cooperation without the Second - Order Free - Rider Problem”, Nature, 432: 499–502.
Pennisi, E., 2005, “How Did Cooperative Behavior Evolve”, Science, 309 (5731): 93.
Porges, S. W., 2011, The Polyvagal Theory: Neurophysiological Foundations of Emotions, Attachment, Communication, and Self - Regulation, New York: W. W. Norton.
Rand, D. G., J. D. Greene, and M. A. Nowak, 2012, “Spontaneous Giving and Calculated Greed”, Nature, 489: 427 - 430.
Rilling, J. K., D. A. Gutman, T. R. Zeh, G. Pagnoni, G. S. Berns, and C. D. Kilts, 2002, “A Neural Basis for Social Cooperation”, Neuron, 35: 395 - 405.
Rilling, J. K., A. G. Sanfey, J. A. Aronson, L. E. Nystrom, and J. D. Cohen, 2004, “Opposing Bold Responses to Reciprocated and Unreciprocated Altruism in Put Rilling, J. K., A. G. Sanfey, J. A. Aronson, L. E. Nystrom, and J. D. Cohen, 2004, “Opposing Bold Responses to Reciprocated and Unreciprocated Altruism in Putative Reward Pathways”, Neuroreport, 15 (16): 2539 - 2543.
Rogers, A. R., 1990, “Group selection by selective emigration: The effects of migration and kin structure”, American Naturalist, 135: 398 - 413.
Rustichini, A., 2005, “Neuroeconomics: Present and Future”, Games and Economic Behavior, 52: 201 - 212.
Sasaki, T. and S. Uchida, 2013, “The Evolution of Cooperation by Social Exclusion”, Proceedings of the Royal Society B, 280: rspb.20122498.
Sethi, R., and E. Somanathan, 1996, “The Evolution of Social Norms in Common Property Eesource Use”, American Economic Review, 86 (4): 766 - 788.
Singer, T., S. J. Kiebel, J. S. Winston, H. Kaube, R. J. Dolan, and C. D. Frith, 2004, “Brain Responses to the Acquired Moral Status of Faces”, Neuron, 41 (4): 653 - 662.
Singer, T., B. Seymour, J. P. O’Doherty, K.E. Stephan, R. J. Dolan, and C.D. Frith, 2006, “Empathic Neural Responses Are Modulated by the Perceived Fairness of Others”, Nature, 439: 466–469.
Trivers, R. L., 1971, “The Evolution of Reciprocal Altruism”, Quarterly Review of Biology, 46: 35 - 57.
Umiltà, M. A., E. Kohler, V. Gallese, L. Fogassi, L. Fadiga, C. Keysers, and G. Rizzolatti, 2001, “I Know What You Are Doing: A Neurophysiological Study”, Neuron, 31: 155 - 165.
Wedekind, C., and M. Milinski, 2000, “Cooperation through Image Scoring in Humans”, Science, 289: 850 - 852.
Wickers, B., C. Keysers, J. Plailly, J. Royet, V. Gallese, and G. Rizzolatti, 2003, “Both of Us Disgusted in My Insula: The Common Neural Basis of Seeing and Feeling Disgust”, Neuron, 40: 655 - 664.
Wiessner, P., 2005, “Norm Enforcement among the Ju/’hoansi Bushmen”, Human Nature, 16 (2): 115 - 145.
Wright, J., 1999, “Altruism as Signal: Zahavi’s Alternative to Kin Selection and Reciprocity”, Journal of Avian Biology, 30 (1): 108 - 115.
Zahavi, A., 1995,“Altruism as a Handicap: The Limitations of Kin Selection and Reciprocity”, Journal of Avian Biology, 26: 1 - 3.
via:
-
人类合作之谜:群体选择及其演化仿真研究
https://html.rhhz.net/NFJJ/html/20221103.htm -
韦倩 姜树广:社会合作秩序何以可能:社会科学的基本问题-山东大学经济研究院
https://www.cer.sdu.edu.cn/info/1032/3450.htm